


REUMATOLOGIA
BÁSICA
PARA
ESTUDANTES
Thelma L Skare
ANO 2019
Faculdade
Evangélica
Mackenzie do PR




























































































Estude fácil! Tem muito documento disponível na Docsity

Ganhe pontos ajudando outros esrudantes ou compre um plano Premium

Prepare-se para as provas
Estude fácil! Tem muito documento disponível na Docsity
Prepare-se para as provas com trabalhos de outros alunos como você, aqui na Docsity
Os melhores documentos à venda: Trabalhos de alunos formados
Prepare-se com as videoaulas e exercícios resolvidos criados a partir da grade da sua Universidade
Responda perguntas de provas passadas e avalie sua preparação.
Ganhe pontos para baixar
Ganhe pontos ajudando outros esrudantes ou compre um plano Premium
Comunidade
Peça ajuda à comunidade e tire suas dúvidas relacionadas ao estudo
Descubra as melhores universidades em seu país de acordo com os usuários da Docsity
Guias grátis
Baixe gratuitamente nossos guias de estudo, métodos para diminuir a ansiedade, dicas de TCC preparadas pelos professores da Docsity
Reumatologia para estudantes, livro bem didático e de fácil compreensão
Tipologia: Resumos
1 / 441

Esta página não é visível na pré-visualização
Não perca as partes importantes!
maneira, cada gene terá que ter o seu linfócito para se expressar. Assim, a população de linfócitos de um indivíduo mostra-se como um verdadeiro mosaico de clones cobrindo todas as possibilidades possíveis de encontro com elementos estranhos. Você já imaginou qual a sua quantidade? Os linfócitos vagueiam pelo organismo em ciclos migratórios exibindo seus receptores como verdadeiras armadilhas. Quando ele encontra o “antígeno da sua vida”, ele se divide formando um clone de células iguais para conseguir fazer frente ao invasor e, assim, dar conta do inimigo. A replicação leva tempo para acontecer, mas pode ser otimizada: num primeiro encontro ela é mais demorada e leva alguns dias para se completar; em encontros subsequentes o processo é bem mais rápido. Trocando em miúdos: quando alguém é exposto a um antígeno, fica melhor protegido contra exposições futuras e este é o fundamento para que se usem vacinas. Um problema que nos interessa de perto e que será objeto de atenção especial mais adiante é o de que as proteínas do próprio corpo podem ser apanhadas nas armadilhas dos linfócitos ocasionando o que chamamos de autoimunidade. Este ataque deve ser prevenido a todo custo, tornando os linfócitos tolerantes aos componentes do próprio corpo. Os mecanismos de tolerância serão discutidos no final deste capítulo.
Bem... Já vimos que a finalidade do sistema imune é livrar-se de elementos estranhos e nocivos ao corpo, que chamaremos a partir de agora, de antígenos. Uma substância para ser reconhecida pelo organismo como antígeno deve ser, antes de tudo, estranha. Isto implica em que o antígeno pode ser um elemento externo ao organismo, ou, um elemento interno alterado, como são as células tumorais e as células infectadas por vírus. No entanto, ser estranho, por si só, não é suficiente. Esta substância deve ter também um tamanho molecular mínimo (quanto maior, melhor) e ter uma certa complexidade química. Quase todos os antígenos são substâncias orgânicas. A incapacidade de substâncias inorgânicas despertarem uma resposta imune talvez se deva ao fato de a maioria ser muito pequena. Moléculas pequenas (como íons metálicos) só podem funcionar como antígeno caso elas se liguem a uma proteína carregadora. Nesta situação elas recebem o nome de hapteno. Outra condição necessária para que uma substância funcione como um antígeno é que ele tenha um grau de complexidade química. Moléculas muito simples, mesmo sendo grandes (como é o caso de certos polímeros que se formam pela repetição de um único aminoácido) são fracamente imunogênicas. Além destes elementos, a concentração em que ela é administrada e a porta de entrada da mesma no organismo, influem na capacidade de uma substância despertar resposta imune ou não. Quando o organismo recebe uma carga muito alta de antígeno ele se torna tolerante a ele; é como se o sistema imune ficasse "atordoado" e perdesse a sua capacidade de reação. Por outro lado, se as concentrações do antígeno são muito baixas, o organismo nem "se incomoda" em responder a ele. Existe, portanto, uma concentração intermediária (que não é nem muito grande, nem muito pequena) capaz de fazer o sistema imune funcionar com capacidade ótima. A via de administração é outro elemento que exerce suas influências e isto é, naturalmente, muito importante quando se leva em conta a administração de vacinas. A administração por via oral acaba sendo raramente utilizada porque as enzimas digestivas destroem o antígeno antes de sua absorção. Aliás, é graças a este mecanismo, que os alimentos normalmente não imunizam uma pessoa. Quando um antígeno é administrado pelas vias subcutânea, intradérmica ou intramuscular acaba atingindo um linfonodo regional onde é processado. Se administrado por via endovenosa ou intraperitoneal, acumula-se predominantemente no baço. Esta diferença de local de processamento talvez seja o responsável por diferenças nas respostas imunológicas.
Como já foi comentado anteriormente, não é a molécula inteira do antígeno que reage com o sistema imune, e, sim, algumas de suas porções chamadas de epítopes ou determinantes antigênicos.
A grande sofisticação do sistema imune dos seres mais evoluídos fica por conta do sistema antecipatório ou antígeno específico cujos agentes efetores são os linfócitos T e B. Quem aprecia um linfócito do ponto de vista morfológico, como células sanguíneas com núcleo denso, e um pequeno anel citoplasmático, sem nenhuma organela especial, a não ser umas poucas mitocôndrias, ribossomos e lisossomos, não imagina que células com uma estrutura tão "vulgar" possam abrigar uma capacidade funcional tão extraordinária. Além disso, essa população, embora sendo anatomicamente homogênea, possui subdivisões bastante distintas do ponto de vista de distribuição de trabalho. A primeira grande divisão dos linfócitos é feita em linfócitos do tipo T e linfócitos B. Os linfócitos T são assim chamados porque, no seu processo de maturação, devem obrigatoriamente passar pelo timo. (T=timo). Os linfócitos B são os linfócitos cuja maturação se processa na medula óssea que seria o órgão equivalente à "bursa de Fabricius" identificada em aves. (B=bursa).
O LINFÓCITO T - Já sabemos que o linfócito T é um linfócito antígeno específico e que a base desta especificidade está na presença de um receptor em sua superfície. A finalidade do receptor é reconhecer um antígeno externo e encaminhar essa informação para dentro da célula para que ela fabrique substâncias que irão ser as verdadeiras efetoras da defesa. Este receptor tem, em sua estrutura básica, 2 cadeias: a α (alfa) e a β (beta). Existem, também, alguns poucos linfócitos que exibem as cadeias γ (gama) e δ (delta), mas só os linfócitos que tem as cadeias αβ pertencem ao sistema imunológico especifico e é deles que iremos falar neste capítulo. Comentaremos sobre os γδ mais tarde mas já fique sabendo que eles são do sistema inespecífico... Cada uma das cadeias do receptor tem uma porção variável (para reconhecer o antígeno) e uma porção constante (que se fixa na superfície celular). A estrutura destas cadeias é geneticamente determinada. Existem genes, melhor falando, famílias de genes para cada uma destas porções. Com isso dá para entender que, se existem grupos de genes para cada uma destas porções, a combinação desta variedade toda resulta num número final muito grande de diferentes receptores. Calcula-se que um único indivíduo pode ter mais de 1.000.000 de clones de células T diferentes... O receptor da célula T não consegue reconhecer o antígeno quando ele está sozinho. Este só passa a ser reconhecido quando for apresentado junto com as moléculas de HLA do próprio indivíduo. De uma maneira geral, o antígeno é processado por uma célula acessória (na maioria das vezes, um macrófago, que, quando exerce esta função, passa a se chamar de célula apresentadora de antígeno) e apresentado ao linfócito T junto com o seu próprio MHC, estimulando-o. Repetindo em outras palavras: o macrófago apanha o antígeno, desmonta-o em pedacinhos menores e expõe todos "os seus segredos" em sua superfície celular junto com o seu antígeno de histocompatibilidade. O linfócito T que encontra o conjunto é, então, ativado. Para que ocorra a estimulação do linfócito T é necessário que ambos, o antígeno e as proteínas do complexo de histocompatibilidade, estejam presentes. Isoladamente, nenhum deles consegue produzir um estímulo efetivo. Existe quem comparasse o comportamento do linfócito T ao de um velho gentleman inglês que segue à risca as regras da etiqueta. Ele sabe que o macrófago expondo o antígeno quer "falar com ele" e sabe até qual é o assunto, mas não irá até ele a menos que seja
citotóxicos. Durante a apresentação do antígeno os linfócitos T helper reconhecem os antígenos apresentados com o HLA classe II e os CD8, os apresentados com HLA classe I. O linfócito T helper pode ser subdividido, mais uma vez, de acordo com as citocinas que ele fabrica, em linfócitos Th1, Th2, Th17 e T regulador (ou T reg). Os linfócitos T CD4+ do grupo Th1 são linfócitos que elaboram citocinas que irão ajudar no combate a bactérias intracelulares. Para entender isto vamos nos lembrar um pouquinho lá da microbiologia.... Lembra que existem certas bactérias, muito “pilantras” que, ao serem fagocitadas continuam crescendo dentro do macrófago? É o que acontece com as bactérias da tuberculose e do mal de Hansen (micobactérias). Isto só é possível porque o vacúolo onde elas estão não consegue se fundir com os lisossomos - que contém as enzimas que deveriam destruí- las. O linfócito Th1 “obriga” os lisossomos do macrófago a irem até o vacúolo onde está a bactéria e despejar o seu conteúdo enzimático. Além disso, estas células Th1 elaboram citocinas que atraem mais macrófagos. Portanto, as células Th1 são fundamentais para a destruição de patógenos intracelulares, ou seja, para a imunidade celular. O segundo subtipo de T helper é o linfócito Th2 o qual auxilia o linfócito B a fabricar anticorpos. Portanto, a célula Th2 está envolvida na resposta humoral. Já a célula Th17 é um linfócito que atua contra micro-organismos extracelulares. A cel T do tipo T reguladora pertence ao sistema de controle da imunidade específica e é uma espécie de “guardinha” que fica atendendo as demais células para que não “aprontem” contra o próprio organismo. Como cada uma dessas células do tipo TCD4 executa essas tarefas? Elaborando determinado tipo de citocinas. Elas são “madames” que não fazem trabalho braçal... Só mandam os outros fazerem através da elaboração de citocinas. Citocinas são substâncias solúveis e de comunicação entre o sistema imune. Como o linfócito T CD4 escolhe qual tipo de célula ele vai se tornar? De novo, por causa das citocinas circulantes... Deu para notar que essas citocinas são realmente “poderosas”! No quadro abaixo estão as principais citocinas que “mandam” na mudança dos linfócitos assim como as principais citocinas que eles fabricam depois que escolheram o seu subtipo... Veja que muitas delas atuam dos dois lados, ou seja... atuam na diferenciação do linfócito e também são elaboradas por ele. Espertinhas, não? Assim o processo faz uma retroalimentação positiva...
FIGURA 1.2 - Subtipos de célula T CD
O linfócito T CD8 ou citotóxico é o que, quando ativado, adquire a capacidade de lisar células alvo, que tem antígenos reconhecidos pelo seu receptor. Ele mata a célula estranha num verdadeiro “abraço de tamanduá”... Chega pertinho dela, modifica o seu exoesqueleto de tal maneira que cria uma área de comunicação com a célula alvo -muitas vezes parecida com um canudinho e, aí, deposita enzimas como granzimas e perforinas que irão lizar esse alvo. Vamos voltar, agora, lá ao encontro do linfócito T com o antígeno de sua vida. Quando ocorre esta ligação começam a acontecer coisas... O primeiro sinal de que existe algo de diferente no ar é um fluxo de Ca++ e Na+ de fora para dentro do linfócito. O interior da célula é normalmente negativo, enquanto que as superfícies externas são carregadas positivamente. Após a ligação com receptor é como se os portões da membrana citoplasmática se abrissem e os cátions fossem despejados de acordo com o gradiente de concentração. À medida que os íons Na+ entram, começam a sair íons H+, o interior da célula se torna basofílico. Outra coisa notada nesta fase precoce é uma mudança na composição dos fosfolipídeos da membrana citoplasmática. Estas alterações iniciais são seguidas por um incremento geral no metabolismo e o que se vê é, em linhas gerais: aumento geral de síntese de proteínas em 2 horas; aumento do número de ribossomos em 6 horas; início da síntese do DNA em 24h. Em 30h o linfócito está pronto para a sua primeira divisão mitótica. Depois desta primeira divisão mitótica, todo o ciclo se repete por 5 a 10 vezes. Feito isto, o linfócito muda novamente: alguns deles retornam para a fase de repouso, outros saem para desempenhar a função imune a que estão destinados. As células que retornam à fase de repouso embora sejam morfologicamente semelhantes à célula-mãe, são diferentes do ponto de vista qualitativo, uma vez que, num segundo encontro com o antígeno elas conseguem responder muito mais ligeiro. São chamadas de células da memória. Já as células que saem para desempenhar a sua função não conseguem mais se proliferar, nem se transformar em células da memória. As que não forem "gastas" ao exercer suas funções morrem em alguns dias após a sua diferenciação. Estas são as células efetoras. As células T da memória conseguem ser encontrada em todos os tecidos do corpo humano, num verdadeiro estado de alerta contra o antígeno. AS células T efetoras são dirigidas para o campode batalha. Já as células T primitivas (ou virgens) só conseguem atravessar o vaso sanguíneo em um ponto especial das vênulas dos órgãos linfoides, onde existem células endoteliais altas. Voltaremos a este assunto ao conversar sobre a circulação dos linfócitos.
O LINFÓCITO B - Outro componente do sistema imunológico específico ou antecipatório é a célula B. Ela é responsável pela formação dos anticorpos ou imunoglobulinas. Da mesma maneira que a célula T, a especificidade da célula B é dada por um receptor de superfície que, neste caso, nada mais é do que uma imunoglobulina de superfície. Esta imunoglobulina de superfície é muito parecida com a imunoglobulina que a célula B secreta como anticorpo, quando estimulada pelo antígeno. Os precursores de célula B têm sua origem na medula óssea e podem ir diretamente aos tecidos linfoides periféricos ou passar pelo fígado, medula óssea, tecido linfoide intestinal ou de amígdala antes de se tornar uma célula funcionante. Um linfócito B, ao se diferenciar da célula tronco da medula óssea, é reconhecido como pré-célula B quando começa a fabricar imunoglobulina M intracitoplasmática. Na fase seguinte ela passa a exibir imunoglobulina M na sua superfície. Depois disso aparece, além da IgM, a IgD de superfície. Tudo que acontece até aqui não depende da presença do antígeno, mas a partir deste estágio a célula já tem a capacidade de responder a elementos estranhos. Se o linfócito B não encontrar o seu alvo, ele morre em 3-4 dias. No entanto, se ele "for salvo" pelo encontro com o antígeno, irá se transformar num linfócito B ativado e irá se diferenciar produzindo uma proporção cada vez maior de imunoglobulina na forma secretada e cada vez menor de imunoglobulina ligada a membrana. Ao encontrar o antígeno algumas destas células B “trocam de roupa” ou seja mudam o tipo de imunoglobulina produzido de IgM para IgA, IgG e IgE mas
Bem... Já sabemos que a finalidade da ativação do linfócito B é a produção de imunoglobulinas que são elementos mediadores da resposta imunológica humoral. Os anticorpos se ligam ao antígeno com a finalidade de inativá-lo ou removê-lo. No homem, existem 5 diferentes classes de imunoglobulinas que são designadas pelas letras: A,G,M,E.D. Cada uma destas imunoglobulinas é formada por 2 cadeias pesadas e 2 cadeias leves. Existem cinco tipos de cadeia pesada (uma para cada tipo de imunoglobulina) e 2 tipos de cadeia leve, de tal modo que são possíveis 10 combinações no total. As cadeias leves (L= light) são chamadas de kappa (k) e lambda (λ), o correspondente, em grego, às iniciais do nome dos seus descobridores. As cadeias pesadas (H=heavy) são identificadas pelas letras gregas correspondentes ao nome das imunoglobulinas, ou seja: gama (γ), para IgG; alfa (α) para IgA; delta (δ) para IgD, mu (μ) para IgM e épsilon (μ) para IgE. As quatro cadeias que formam a imunoglobulina se ligam entre si por pontes dissulfetos. Cada uma das cadeias tem uma porção variável (V) e uma porção constante (C). Assim, as cadeias leves têm uma porção variável (VL) e uma porção constante (CL); as cadeias pesadas também têm uma porção variável (VH) e uma constante, (CH). Como o nome sugere, a porção constante é feita da mesma estrutura primária nas imunoglobulinas do mesmo tipo e subclasse. A porção constante está envolvida nas atividades biológicas das imunoglobulinas. Ex: CH2 da IgG e CH4 da IgM estão envolvidas na ligação com o complemento. Isso esta muitobem feito, porque várias células que fazem fagocitose têm receptores para estas porções - o que facilita o trabalho. Então elas não podem ficar variando muito, por ai... As porções variáveis (VL e VH) constituem a região de ligação ao antígeno. Dentro da porção variável existe uma porção hipervariável chamada idiotipo que é única para cada molécula de imunoglobulina. Anticorpos contra esta porção são chamados de anticorpos anti- idiotípicos e ocorrem normalmente durante a resposta imune. Parece que eles geram um sinal para a célula B "desligar" a sua produção de anticorpos. Quando uma molécula de imunoglobulina é submetida à digestão por papaína (que age na região entre CH1 e CH2 conhecida como dobradiça), resultam 2 fragmentos: o Fab ( a = antigen, b = binding ) e o Fc (c = crystallizable , mas este c pode também servir para C omplemento uma vez que é esta a porção que se liga ao complemento ou C élula- porque também se liga aos receptores de fagócitos já mencionados).
FIGURA 1.4 - Estrutura da Imunoglobulina
As principais características das imunoglobulinas estão resumidas no quadro 1.1.
Isotipo Cadeia pesada Cadeia leve Ativação do complemento clássica alternada IgG (^) 1 2 3 4 e IgG 1,2 e 3 Ig G IgM e + - IgA (^) 1 2 e - + IgD e - + IgE (^) e - +
As imunoglobulinas G formam aproximadamente 75% das imunoglobulinas totais do soro. As classes de IgG, em número de 4 (IgG1; IgG2; IgG3; IgG4) estão numeradas em ordem decrescente de concentração sérica, sendo a IgG1 encontrada em maior concentração e a IgG em menor. Esta imunoglobulina atravessa a barreira placentária e proporciona uma linha importante de defesa contra infecções durante os primeiros meses de vida do recém-nascido. Anticorpos do tipo IgG são ótimos no preparo de células e partículas para ingestão pelos fagócitos, ou seja ,são excelentes opsoninas. Neste caso, a IgG se liga pela porção Fab ao antígeno e pela porção Fc a um receptor especial para Fc de IgG que existe nos fagócitos. A seguir todo o conjunto é engolfado pelo fagócito. A IgG é um anticorpo de resposta secundária, ou seja, quando ocorre uma estimulação antigênica formam-se primeiro anticorpos tipo IgMs (que são os anticorpos de resposta primária) e mais tarde IgGs. A maioria dos subtipos de IgG ativa complemento pela via clássica. A imunoglobulina M circula como um pentâmero, ou seja, como um conjunto de 5 monômeros ligados entre si pela cadeia J. É, como já foi dito anteriormente, um anticorpo de resposta primária, ou seja, é o primeiro anticorpo a aparecer no organismo após uma infecção. É, também, o primeiro anticorpo a ser fabricado pelo recém-nato. A IgM, graças ao fato de ser um pentâmero e portanto, com muitos pontos de ligação com o antígeno, é eficiente nas reações de aglutinação. Além disso, o seu tamanho grande faz com que ela tenha tendência para permanecer no espaço intravascular, e, não consiga atravessar a barreira placentária. Uma das suas principais maneira de agir é através de fixação do complemento, promovendo lise do antígeno. A imunoglobulina A é responsável por 10 a 15% das imunoglobulinas do soro, mas é a principal imunoglobulina das secreções (saliva, lágrima, secreções nasais e gastrintestinais e leite humano). É especialista em se transportar para áreas do corpo inacessíveis às células produtoras de anticorpos. Nas secreções a IgA está em sua forma secretória (sIgA), que é um polímero contendo geralmente dois monômeros, uma peça de junção (cadeia J) e uma glicoproteína chamada de peça secretória. A IgA fixa complemento via alternativa e tem uma ação antiviral importante por prevenir a ligação do vírus com as células epiteliais do aparelho respiratório e gastrointestinal. Ela funciona como uma "cola" grudando os agentes patógenos todos juntos e impedindo a sua penetração no organismo. Existem duas subclasses de IgA. A IgA1 que é a principal IgA do soro e a IgA2 que é a prevalente nas secreções. Esta imunoglobulina é transportada pela placenta para o feto e também secretada pelo leite materno. A imunoglobulina D é encontrada em concentrações bem baixas e junto com a IgM é o maior receptor de antígeno de superfície de linfócito B. A imunoglobulina E também é encontrada em baixas concentrações. Estas Igs têm receptores especiais em mastócitos e basófilos de tal maneira que, quando ocorre a ligação IgE com antígenos, estas células liberam o conteúdo de seus grânulos, que são substâncias que vão promover contratura de musculatura lisa e assim tentar a expulsão de parasitas. Infelizmente, muitas vezes estas IgE reagem com elementos do meio ambiente como, p. ex., pólen, poeira, etc e a consequente liberação de material de mastócitos e basófilos é responsável pelo aparecimento de reações alérgicas.
Como já comentado anteriormente interleucinas ou citocinas são substâncias que servem para a comunicação celular. As células interligadas por estes agentes devem ter receptores especiais para a citocina em questão e podem pertencer tanto ao sistema imunológico especifico como inespecífico ou, ainda, serem células que não pertençam ao sistema imunológico. Quimiocinas, como o nome sugere, atuam como elementos quimiotáticos sendo responsável pela movimentação celular. Estas substâncias têm se tornado alvos importantes no tratamento das doenças reumáticas e existem fármacos que conseguem inibí-las ou ocupar o seu receptor. Por isso, embora pareça meio chato, é necessário que você guarde as principais. O grupo mais importante das citocinas pró-inflamatórias é a tríade IL-1, IL-6 e TNFα e essas três podem ser bem inflamatórias quando estão sozinhas... Imagine só quando se juntam... Na reumatologia existem medicamentos que inibem o TNFα (infliximabe, etanercepte, golimumabe, adalimumabe e certolizumabe); que inibem a IL-6 ( tocilizumabe) e a IL-1 ( anakinra e canaquinumabe). Apesar dos nomes horrorosos eles são ótimos anti-inflamtorios e imunomoduladores e têm mudado a historia natural de muitas doenças reumáticas. Aqui vão as principais citocinas:
Interleucina-1 (IL-1): É responsável pela indução da saída de neutrófilos da medula óssea, e, uma vez que eles estejam circulando, promove a sua marginalização e entrada nos tecidos. É, portanto, uma das grandes atrizes do processo inflamatório. Em músculo, promove liberação de prostaglandinas, estimula proteólise e liberação de aminoácidos o que, a longo prazo, resulta em perda de massa muscular. Em sistema nervoso central promove sonolência, anorexia e pode agir como pirógeno. Linfócitos T e B também sofrem ação da IL-1. Neles esta linfocina induz diferenciação e proliferação, aumenta produção de IL-2, atividade de célula T citotóxica e da natural killer. A ação da IL-1 está na dependência de sua ligação com receptores específicos. Existe, também, um receptor “falso” para a IL-1, ou seja, um receptor que se liga com a sua molécula, mas não passa a informação para a frente, funcionando como um bloqueador. (Faz inibição competitiva.) Este tipo de receptor é chamado de antagonista de receptor de IL-1 (IL-1-Ra).
Interleucina-6 (IL-6): Age em estádios finais de diferenciação de célula B e induz a formação de reagentes de fase aguda pelos hepatócitos erguendo as provas de atividade inflamatória como VHS e proteína C reativa. Ativa proliferação de célula B e T, síntese de imunoglobulina e medeia alguns dos efeitos da IL-1. A inibição da IL-6 é usada no tratamento da artrite reumatoide do adulto e é feita através do tocilizumabe.
Fator tumoral de necrose (TNF): é um grupo de substâncias produzidas principalmente por macrófagos ativados e que promove uma necrose hemorrágica em células tumorais (daí o seu nome). É ela o elemento responsável direta ou indiretamente (através de liberação de IL-1) pela febre, recrutamento e ativação dos neutrófilos e monócitos em infecções. O TNF α também é essencial para a formação de granulomas como os que atuam na defesa contra as micobactérias. Junto com IL-6 atua no fígado levando à formação dos reagentes de fase aguda. Produção prolongada de TNF α causa perda de massa muscular e de tecido adiposo ocasionado a caquexia. Em casos de sepsis por bactérias gram negativas existe produção maciça de TNFα que é o responsável por alguns dos achados do choque séptico
Interleucina 10: Tem atividade anti-inflamatória porque inibe macrófagos e células dendriticas ativadas.
Interlecina 4: Estimula a formação de IgE e suprime reações que dependem de macrófagos.
Transforming Growth Factor β (TGF-β): Inibe a proliferação e ativação de linfócitos e outros leucócitos.
Interferons (INF) tipo I: Os principais são os INF α e INFβ. O nome interferon vem da capacidade que estas substâncias têm de interferir na infecção viral que é a sua principal finalidade.
Interferon tipo II : É o INF γ. Ativa macrófagos aumentando a sua capacidade microbicida; atua na troca de fabricação de tipo de imunoglobulinas pelo linfócito B e diferencia células T em Th1.
Interleucina 2: É elaborada pelo linfócito T e é um grande fator para o seu próprio crescimento respondendo pela sua expansão clonal após o reconhecimento do antígeno
Interleucina 17 : Tem atuação pró-inflamatória e está associada a processos alérgicos. Induz a formação de IL-6, GM-CSF, IL-1β, TGF-β, TNF-α, quimiocinas e prostaglandinas.
Existe um grande número de outras citocinas que não serão abordadas aqui, porque isto escapa ao intuito desta revisão. Para guardar o principal veja que IL-1 IL-6 e TNFα são pró- inflamatórias e que a IL-10, IL-4 e TGFβ tem atividade anti-inflamatória. Quando uma citocina se liga ao seu receptor que está na membrana da célula, ela tem que mandar a mensagem lá para dentro da célula para que, de uma forma ou outra, chegue ao núcleo e o DNA “tome as devidas providências” para cumprir a ordem da citocina. Para isso ela vai precisar de “mensageiros” que travessem o citoplasma e cheguem ao núcleo. Por que saber isso? Porque existem alguns medicamentos que podem inibir alguns esses mensageiros e assim anular a ordem da citocina. Um desses intermediários é um sistema de enzimas chamado de JANUS kinases (ou JAKs - para os mais íntimos). Outro grupo é o dos STATs ( Systems Transducers and Activators of Transcription ). Os sistemas JAK/STATs podem ser usados por mais do que uma citocina. Assim é possível, inibindo só um deles, impedir a atuação de várias citocinas ao mesmo tempo. Um aparte curioso: o nome Janus das nossas amigas JAKs vem do deus romano JANUS que deu origem ao nome do mês de janeiro e que era o deus das portas (das entradas, saídas e das transições). Ele era representado por uma figura masculina com duas faces: cada uma olhando para um lado: o de dentro e o de fora da porta... A “nossa” JAK olha para a ordem da citocina que se ligou ao receptor e para as reações que irão ao núcleo aonde se cumprirão as ordens da dita.
Microbiologistas descobriram no final do século 19 que existiam algumas substâncias que deveriam estar presentes no soro de um ser vivo para que os elementos estranhos pudessem ser efetivamente combatidos. Uma delas - que era estável ao calor - foi chamada de complemento porque complementava a ação bactericida do anticorpo. Descobriu-se, depois, que o complemento não era uma substância única e, sim, um verdadeiro time, com pelo menos 20 jogadores. Normalmente estes componentes estão na forma inativa, mas, a um sinal específico, o primeiro componente do time se ativa e começa todo o processo, o qual, segundo um imunologista tem um comportamento muito parecido com o de corridas de bastão. Uma vez ativado o primeiro componente, este vai até o segundo e "passa o bastão". Este segundo componente se ativa e corre até o terceiro "passando-lhe o bastão" e assim por diante. O último membro do time termina a corrida fazendo um buraco na membrana da célula invasora através da formação do complexo de ataque à membrana.
Os elementos capazes de ativar cada uma dessas vias são diferentes. A via clássica é ativada por complexos imunes contendo IgG1, IgG2, IgG3 e IgM e também por diversas substâncias químicas como proteína C reativa, DNA e enzimas semelhantes à tripsina. A via alternativa é ativada por complexos imunes contendo certos polissacarídeos como endotoxinas bacterianas e cápsulas de fungos, membranas de diálise, veneno de cobra e contrastes endovenosos usados para estudos radiológicos. A via da manose-lectina é ativada por certos antígenos que têm moléculas com resíduos de manose e outros açúcares que seguem uma determinada orientação espacial e são encontrados em vários micro-organismos. Todas as vias convergem para a ativação do componente C3, que é onde se inicia a via terminal ou comum. Eventualmente a via comum ou terminal pode ser ativada diretamente por certas enzimas que não fazem parte do complemento, sem a participação das vias iniciais de ativação. Será que agente precisa mesmo saber todos os componenetes do complemento? Não Apenas os principais. Saiba que na via clássica estão C1, C2 e C4. E que o C3 está no início da comum. Isto é o suficiente para o dia a dia.
FRAGMENTOS DE COMPLEMENTO - Além do complexo de ataque à membrana que é o produto principal e final da ativação do complemento, vários outros fragmentos formados durante a cascata de ativação têm atividade biológica. Veja, no quadro 1.3, os principais - os quais podem ser resumidos como: C3a e C5a são anafilatocinas e o C3b é opsonina.
C3a: - anafilatoxina, libera histamina dos mastócitos; C3b :- funciona como opsonina, ou seja, favorece a aderência do complexo-imune, o qual é um prelúdio à fagocitose. Além disso, várias células (eritrócitos, PMNs, e linfócitos) contêm receptores para este fragmento. Estes receptores ajudam no transporte do complexo imune até órgãos do SRE onde serão metabolizados. Na verdade, eles funcionam como verdadeiros ganchos para o complexo imune que assim se liga às células circulantes e vai de "carona" até órgãos do SRE. O número destes receptores é altamente variável na população e significativamente reduzido em pacientes com lúpus; C5a:- anafilatoxina, libera histamina dos mastócitos, quimiotático para polimorfo nucleares e monócitos. Aumenta a produção de moléculas de adesão.
MECANISMOS DE CONTROLE DO SISTEMA DO COMPLEMENTO - A ativação não controlada do sistema imune é prevenida pela labilidade dos sítios ativados, e, pela dissociação da C convertase dependente de tempo e temperatura. Existem também algumas proteínas séricas que servem para modular e limitar esta ativação, tais como: inibidor de C1; fator I e o fator H.
DOSAGEM DO COMPLEMENTO - Medir o complemento pode ser útil no diagnóstico e avaliação do grau de atividade de uma doença assim como, da resposta ao tratamento. O resultado da dosagem do complemento reflete o balanço entre o complemento sintetizado e aquilo que é consumido. Deve-se lembrar que, no entanto, em certas condições inflamatórias, frações do complemento têm comportamento de reagentes de fase aguda, podendo, antes de se iniciar o consumo, ter um nível aumentado. Com isto o valor final após o consumo simplesmente "cai para um valor normal", podendo dar uma impressão falsa, de que não existe consumo. Além disso, situações de síntese diminuída, como é a desnutrição severa e a insuficiência hepática (porque é lá que são fabricados os componentes do complemento), promovem um valor final baixo, sem que exista consumo.
No quadro 1.4, algumas das situações associadas com o achado de uma dosagem diminuida de complemento.
HIPOSSÍNTESE: deficiências congênitas, insuficiência hepática,desnutrição HIPERCATABOLISMO: deficiência de proteínas controles: Ex: deficiências de C1 inibidor; doenças reumáticas com depósito de complexo-imune : LES, vasculites AR. doenças infecciosas : endocardite, hepatite B, sepsis etc doenças renais: Ex : glomerulonefrites pós-estreptocócica, membrano-proliferativa e proliferativa idiopática.
Outro aspecto a ser lembrado é a alta frequência de deficiências congênitas de síntese do complemento, as quais podem ser causa de uma dosagem sérica baixa. Algumas pessoas com defeito de complemento são normais; outras têm, no entanto, maior incidência de certas doenças como, por exemplo, o lúpus. Veja alguns exemplos no quadro 1.5. Note como as deficiências dos componentes iniciais se associam com doenças reumáticas; as demais com doenças infecciosas.
C1q:- glomerulonefrite, poiquiloderma congênito C1r:- glomerulonefrite, lúpus - like C1s:- lúpus like C4:- LES, e síndrome de Sjögren C2:- LES, lúpus discóide, Hodgkin e glomerulonefrites
C3:- lúpus - like e glomerulonefrites C5:- LES, infecções por Neisseria C6:- Infecções por Neisseria C7:- LES, AR e infecções por Neisseria C8:- LES, infecções por Neisseria C9:- infecção por Neisseria
Embora os defeitos homozigóticos sejam relativamente raros, os defeitos heterozigóticos são relativamente comuns. Quando se pretende avaliar a atividade de uma doença através daquilo que ela está consumindo de complemento, pode-se proceder à medida do complemento hemolítico total (CH50) ou dos componentes individuais (os mais usados são C3 e C4). A dosagem do complemento hemolítico total representa o resultado da atividade de todos os componentes do sistema, atuando de maneira encadeada, sem especificar se o processo começou via clássica ou via alternativa. A dosagem de C3 também está diminuída em casos de ativação de ambos os braços do complemento, visto que este elemento se situa na via comum. Já o C4, só estará diminuído em casos de ativação da via clássica e da do receptor de manose-lectina. Repetindo: uma diminuição de C3, C4 e CH50 indica que está existindo consumo do complemento via clássica (Ex: LES); uma diminuição de C3 e CH50 com C4 normal indica que este consumo ocorre em via alternada (Ex: glomerulonefrite, choque endotóxico). Uma medida de atividade CH50 de zero aponta para deficiência hereditária total de um dos componentes, o que interrompe a sequência dos eventos totalmente, impedindo que se forme o complexo de ataque à membrana.
Bem, já analisamos a parte mais sofisticada do sistema imune que é privilégio dos animais mais evoluídos. Só que, enquanto ela não age com toda a sua potência, o organismo fica na dependência de uma defesa não específica, que nem por isso é menos importante e
A fase da digestão... Depois que a partícula se fixa na membrana do fagócito ela é engolfada formando uma vesícula, o fagossomo, que nada mais é do que uma bolsa formada ao redor do elemento estranho, às custas de membrana citoplasmática. O citoplasma dos fagócitos está repleto de pequenos vacúolos que estão cheios de enzimas. Esses vacúolos se chamam grânulos nos neutrófilos e lisossomos nos macrófagos; a principal diferença entre elas está no seu conteúdo enzimático. Os grânulos e os lisossomos estão se movendo livremente e sem direção definida dentro da célula. Assim que o vacúolo fagocítico começa a se formar estes grânulos colidem com o fagossomo e se aderem a ele despejando todo o seu conteúdo (e formando um fagolisossomo). De verdade, o esvaziamento do conteúdo enzimático para dentro do fagossomo começa quando ele ainda não se fechou completamente e pode existir escape de enzimas para o meio extracelular, naturalmente, com todo o potencial de lesão do mesmo. Uma das consequências da degranulação é a acidificação do fagossomo, a qual facilita a morte do organismo ingerido. Além da bactéria não conseguir sobreviver em meio ácido, muitas das enzimas funcionam melhor neste meio.
O que fazer com as sobras... Sempre que o fagócito destrói o elemento invasor aproveita muito da sua matéria prima, como por exemplo, aminoácidos, açúcares e nucleotídeos, os quais são reutilizados no metabolismo da célula. O problema é com os restinhos indigeríveis. Para eles, o fagócito tem 3 opções possíveis, que são: a defecação, a indigestão ou a morte. Se ele optar pela primeira possibilidade, irá despejar no meio extracelular não só os restos indesejáveis como também muitas enzimas potencialmente danosas. A segunda alternativa leva o fagócito a estocar material indigerível dentro dos lisossomos, os quais se fundem e formam uma grande vesícula cheia de lixo. Optando por esta possibilidade o fagócito fica "cronicamente constipado" e isto também não é bom para o organismo. Na última opção, o fagócito acumula o material indesejável em tantos quantos vacúolos ele conseguir e, após isto, ele morre. O "cadáver" é expelido do corpo na forma de secreções purulentas etc... Esta última forma de opção parece ser a mais adequada e a mais frequente. Quando uma célula exerce a fagocitose, segue-se um pico de consumo de oxigênio por esta célula. Este consumo de oxigênio não tem nada a ver com o processo respiratório o qual é executado pelas mitocôndrias. Na realidade, ele depende de um conjunto de enzimas - o complexo NADPH oxidase, que existe na membrana da célula (e também na membrana do
fagossomo, a qual é a própria membrana da célula invaginada). A finalidade deste conjunto de reações é converter oxigênio molecular em ânion superóxido, peróxidos, oxigênio singlet e radicais hidroxilas. Destes, os 3 primeiros representam radicais de oxigênio livre; o radical hidroxila não é um radical de oxigênio livre mas também é altamente reativo. Devido a este alto grau de reatividade, estas substâncias são altamente tóxicas, atacando as partículas que são ingeridas. O ânion superóxido, por ser uma substância altamente instável, não tem tempo de participar deste ataque, mas se transforma em outros radicais de oxigênio livre que serão os elementos ativos. O peróxido de hidrogênio é um agente germicida bem conhecido e sua atividade aumenta ainda mais quando reage com agentes halogenados. Da interação com cloretos, por exemplo, resulta a formação do ácido hipocloroso que é, também, agente bactericida. Ao contrário dos elementos oxidantes, os quais têm vida curta e são incapazes de selecionar seu alvo, as enzimas proteolíticas contidas nos grânulos permanecem ativas por um longo período de tempo e se dirigem para um substrato selecionado, podendo catalisar uma mesma reação diversas vezes. Os grânulos dos neutrófilos contem um grande número de enzimas, porém 3 delas nos interessam de perto pelo grande potencial de destruição tissular: a elastase e duas metaloproteinases (a colagenase e a gelatinase). Não deixa de ser interessante notar o fato de que enzimas como colagenases e gelatinases são liberadas como pró-enzimas (inativas) e necessitam de exposição aos radicais de oxigênio livre para a sua ativação. Os grânulos dos eosinófilos têm várias enzimas com propriedades anti-inflamatórias como histaminase, arilsulfatase e fosfolipase D e, por isso estas células parecem desempenhar um fator regulador no término do processo inflamatório. Quanto aos basófilos, as suas funções não são bem compreendidas. Acredita-se que mediadores liberados pelos basófilos aumentem a permeabilidade vascular e assim melhore a entrega de outros elementos da resposta imune. Os basófilos têm em sua superfície receptores para IgE, C3a e C5a. A ligação de receptores para IgE com a imunoglobulina e a ligação desta com o antígeno pode levar a liberação de seus grânulos que são ricos em histamina, proteases neutras e mediadores da resposta de hipersensibilidade imediata (anafilaxia).
A NETOSE – Embora a função mais conhecida de um neutrófilo seja a de eliminar o elemento estranho por digestão enzimática, descobriu-se recentemente que estas células são, na verdade, muito mais sofisticadas do que parecem à primeira vista. Uma dessas descobertas mostra que o neutrófilo é capaz de eliminar microorganismos, formando uma rede com a sua própria cromatina (DNA + histona) misturada com suas enzimas lisossômicas. A cromatina pode se originar do seu núcleo ou das mitocôndrias. Esta rede tem um aspecto de “teia de aranha” na qual os microorganismos ficam presos e são eliminados. Isto acontece mais comumente com um tipo de neutrófilo menorzinho chamado de neutrófilo de baixa densidade. Nesta brincadeira o PMN pode ou não morrer. Já a exposição do material intranuclear e das enzimas lisossômicas ao meio extracelular - que é um material que mormalmente fica escondido dentro da célula - pode parecer “novidade“ ao sistema imune e despertar autoimunidade. Pacientes com lúpus parecem ser mais propendos a ter PMNs que façam netose.
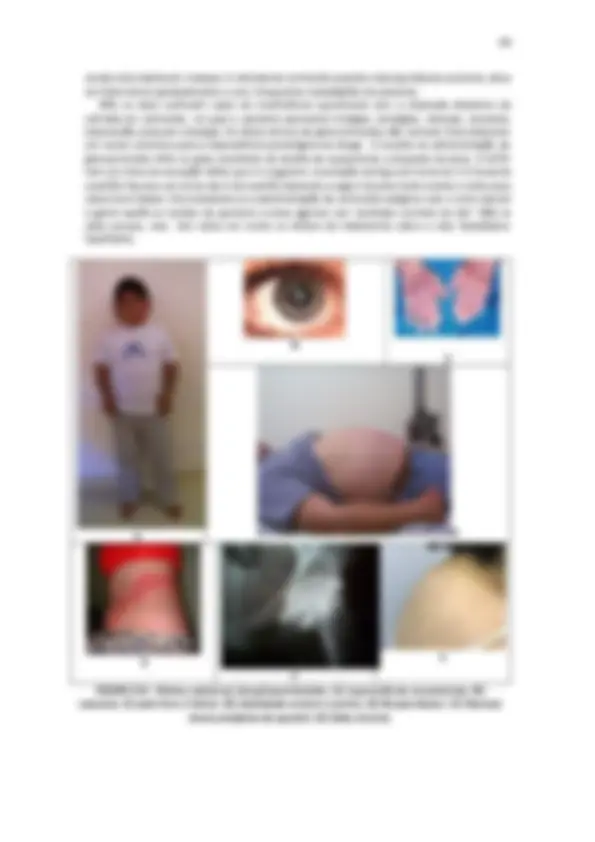
FIGURA 1.6 - Mecanismo esquemático da netose