Download Aquatic Adaptations in Spinosaurus: Departure from Land-based Theropods and more Lecture notes Biology in PDF only on Docsity!
Tail-propelled aquatic locomotion in a theropod dinosaur
Authors: Nizar Ibrahim* 1 , Simone Maganuco2,3, Cristiano Dal Sasso 4 , Matteo Fabbri 5 , Marco Auditore 6 , Gabriele Bindellini7,6, David M. Martill 8 , Samir Zouhri 9 , Diego Mattarelli 6 , David M. Unwin 10 , Jasmina Wiemann 5 , Davide Bonadonna 3 , Ayoub Amane 9 , Juliana Jakubczak 1 , Ulrich Joger 11 , George V. Lauder 12 , Stephanie E. Pierce* 12 Affiliations: 1 Department of Biology, University of Detroit Mercy, Detroit, MI 48221, USA. 2 Associazione Paleontologica Paleoartistica Italiana, 43121 Parma, Italy. 3 Associate of 4. 4 Sezione di Paleontologia dei Vertebrati, Museo di Storia Naturale di Milano, 20121 Milan, Italy. 5 Department of Geology & Geophysics, Yale University, New Haven, CT 06511, USA. 6 Collaborator of 4. 7 Dipartimento di Scienze della Terra “A. Desio”, Università degli Studi di Milano, 20133 Milan, Italy. 8 School of the Environment, Geography and Geological Sciences, University of Portsmouth, Portsmouth, PO1 3QL, UK. 9 Department of Geology, Faculty of Sciences Aïn Chock, Hassan II University of Casablanca, Morocco. 10 School of Museum Studies, University of Leicester, Leicester LE1 7RF, United Kingdom
11 Staatlich Naturhistorisches Museum Braunschweig, 38106 Braunschweig, Germany. 12 Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology, Harvard University, Cambridge, MA 02138, USA. *Correspondence to: ibrahini@udmercy.edu and spierce@oeb.harbard.edu Intensive research on non-avian dinosaurs in recent decades strongly suggests that they were restricted to terrestrial environments 1. Historical views proposing that some groups, such as sauropods and hadrosaurs, lived in aquatic environments2,3 were abandoned decades ago4,5,6. Recently, however, it has been argued that at least some spinosaurs, an unusual group of large-bodied Cretaceous theropods, were semi-aquatic7,8, but this idea has been challenged on anatomical, biomechanical, and taphonomic grounds and remains controversial9,10,11. Here we present the first unambiguous evidence for an aquatic propulsive structure in a dinosaur, the giant theropod Spinosaurus aegyptiacus 7 , 12****. This dinosaur has a tail with an unexpected and unique shape consisting of extremely tall neural spines and elongate chevrons forming a large, flexible, fin-like organ capable of extensive lateral excursion. Using a robotic flapping apparatus to measure undulatory forces in physical tail models, we show that the tail shape of Spinosaurus produces greater thrust and efficiency in water than the tail shapes of terrestrial dinosaurs, comparable to that of extant aquatic vertebrates that use vertically expanded tails to generate forward propulsion while swimming. This conclusion is consistent with a suite of adaptations for an aquatic lifestyle and a piscivorous diet in Spinosaurus 7 ,13,14****. Although developed to a lesser degree, similar aquatic adaptations are found in other spinosaurids15,16 – a clade with a near global distribution and a stratigraphic range of more than 30 million years 14 documenting a persistent and significant invasion of aquatic environments by dinosaurs.
Here we describe a nearly complete, partially articulated tail of a subadult individual of Spinosaurus aegyptiacus ( FSAC-KK 11888), from the Cretaceous Kem Kem beds of south- eastern Morocco (Figs. 1, 2, Extended Data Figs. 1-4, Supplementary Information Part 1, Supplemental Information Video 1). The skeleton represents the most complete dinosaur known from the Kem Kem21,22 and the most complete skeleton of a Cretaceous theropod from mainland Africa (Supplemental Information Part 2). As we show here, the tail forms part of the neotype of S. aegyptiacus 7 and was found in direct juxtaposition to the remainder of the skeleton (Extended Data Fig. 3). Over 90% of the new material, which confirms that a single subadult individual is preserved at the site, was recovered during field excavations in late 2018 and digitally recorded (Extended Data Fig. 5, Supplementary Information Parts 2-4). Several elements compare closely to drawings of Spinosaurus fossils destroyed in World War II (Extended Data Fig. 6). More than 30 near-sequential caudal vertebrae (located within caudal positions 1-41) of FSAC- KK 11888 are preserved, representing approximately 80% of original tail length (Extended Data Figs. 3, 4, Extended Data Table 1). Both proximal and distal elements of the tail are complete and preserved in three-dimensions, indicating minimal taphonomic distortion (Fig. 2, Supplemental Information Video 2). At the level of the caudal transition point 1 , the centra become proportionally more elongate. In addition, the prezygapophyses no longer overhang the preceding centrum and show a marked decrease in size compared to many theropod dinosaurs 1. The postzygapophyses also decrease in size, leading to a reduced contact with the prezygapophyses, and are completely absent in the distalmost caudal vertebrae (Fig. 2). This again is different from the condition seen in most theropods, where zygapophyses become more elongate and more prominent toward the tail tip 1 restricting flexibility in more distal intervertebral joints.
The neural arches are distinctive elements of the Spinosaurus tail. A remarkable complex of vertebral laminae and fossae is present in the proximal caudal vertebrae, and partly persists in mid-caudal neural arches. The morphology of the neural spines shows considerable variation along the sequence (Figs. 1, 2, Extended Data Table 1): spines of proximal caudals are about three times taller than their centra and are cross-shaped in cross-section from base to mid-height, in mid-caudals the spines become much longer, and in the small distal caudals neural spine length reaches well over seven times the height of the centrum (contra ref. 11 ). The neural spines of mid-distal caudal vertebrae of Spinosaurus have a unique cross-section, whereby they are proximo-distally rather than mediolaterally flattened. This is due to hyper-developed spinodiapophyseal laminae and loss of pre- and postspinal laminae. The tail chevrons also differ from those of other theropods. Their morphology varies little throughout the caudal series, except for a slight gradual reduction of the haemal canal: distal chevrons are as elongate as the proximal ones, but they become slender, paralleling the gradual decrease in size of the centra. Taken together, the elongated neural and haemal arches result in a dramatic, vertically expanded tail shape with extensive lateral surface area (Supplemental Information Fig. 4). The skeletal anatomy of Spinosaurus represents a major departure from that of other theropods including the clade within which it is located 1 , the basal Tetanurae, which comprises crown group birds and all other stem theropods more closely related to birds than to Ceratosaurus 1. A key feature of this group is a stiffened tail in which the degree of overlap in articulation between pre- and postzygapophyses increases along the caudal series, drastically diminishing the range of motion between individual vertebrae 1. This trend in motion reduction is emphasised in paravians with the appearance of ossified ligaments and/or reduction and fusion of the caudals into a pygostyle 17. By contrast, in Spinosaurus the pre- and postzygapophyses are much further reduced
narrow tails of terrestrial theropods, supporting the inference that Spinosaurus utilized tail propelled swimming. This tail morphology may have also increased lateral stability of the body in the water, reducing the tendency to roll while floating 10. Contrary to recent suggestions 10 that Spinosaurus was confined to wading and the apprehension of prey from around the edges of water bodies, the morphology and function of the tail along with other adaptations for life in water 7 point to an active, highly specialized aquatic predator that pursued and caught its prey in the water column (Extended Data Fig. 7). The skeletal remains of Spinosaurus (SI) from the Kem Kem beds – composed of sediments deposited in a major fluvio- deltaic system 7 that have yielded a diverse vertebrate assemblage 27 – provide further insights into the ecology of this dinosaur. The Kem Kem assemblage is highly atypical, containing a rich freshwater fauna dominated by fish, including lungfish and large to very large sawfish, and coelacanths 27 , a diverse range of crocodyliforms 28 , and several giant predatory dinosaurs7,22. The seemingly anomalous occurrence in the same deposits of several large-bodied predators, but few terrestrial herbivores, is partially explained by the fully aquatic, likely piscivorous, lifestyle of Spinosaurus which considerably expands the morphological and ecological disparity of Kem Kem tetrapods7, 29. At the same time, competition with several co-occurring large aquatic predators 28 may have driven the evolution of giant size in Spinosaurus. The discovery that an evolutionarily significant clade of dinosaurs exploited environments previously thought to be uninhabited by this group highlights weaknesses in current datasets upon which global evolutionary narratives are based. Dinosaur anatomy and functional morphology are perhaps rather less well understood than often claimed 30. This is likely related, in part, to the under- representation of dinosaur remains from Africa, the sampling of which lags far behind that of Europe, Asia and North America 1. Other African dinosaurs preserved in similar settings, such as
the poorly understood “hippopotamus ornithopod” Lurdusaurus 31 , may also have been better adapted to aquatic settings than currently recognized, further emphasising current limitations to our comprehension of these animals. References
- Weishampel D. B., Dodson P., & Osmólska H. The Dinosauria 2nd edn. (Berkeley, Univ. of California Press, 2004).
- Owen, R. A description of a portion of the skeleton of the Cetiosaurus , a gigantic extinct saurian reptile occurring in the oolitic formations of different portions of England. Proceedings of the Geological Society of London 3 , 457-462 (1841).
- Cope, E. On the characters of the skull in the Hadrosauridae. Proceedings of the National Academy of Sciences of Philadelphia 35 , 97-107 (1883).
- Kermack, K. A. A note on the habits of sauropods. Annals and Magazine of Natural History 12 , 830-832 (1951).
- Bakker, R. T. Ecology of the brontosaurs. Nature 229, 172 - 174 (1971).
- Alexander, R. McN. Mechanics of posture and gait of some large dinosaurs. Zool. J. Linn. Soc. , 83 , 1–25 (1985).
- Ibrahim, N. et al. Semiaquatic adaptation in a giant predatory dinosaur. Science 345 , 1613- 1616 (2014).
- Aureliano, T. et al. Semi-aquatic adaptations in a spinosaur from the Lower Cretaceous of Brazil. Cretaceous Res. 90 , 283-285 (2018).
- Witmer, L. M. The debate on avian ancestry: Phylogeny, function, and fossils. in Mesozoic Birds: Above the Heads of Dinosaurs (eds. Chiappe, L. M. & Witmer, L. M.) 3–30 (Univ. California Press, 2002).
- Tereshchenko, V. Adaptive features of protoceratopoids (Ornithischia, Neoceratopsia). Paleontol. Journal 42 , 273-286 (2008).
- Cau, A. et al. Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs. Nature 552 , 395– 399 (2017).
- Sereno, P. C. et al. A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids. Science 282 , 1298-1302 (1998).
- Lavocat, R., Comptes rendus 19e Congrés géologique international (Alg.) 1952. Acad. Sci. Paris 15 , 65 (1954).
- Sereno, P. C., Dutheil, D. B., Iarochene, M. & Larsson, H. C. Predatory dinosaur from the Sahara and Late Cretaceous faunal differentiation. Science 272 , 986-991 (1996).
- D'Août, K. & Aerts, P. Kinematic and swimming efficiency of steady swimming in adult axolotls ( Ambystoma mexicanum ). J. Exp. Biol. 200 , 1863-1871 (1997).
- Fish, F. Kinematics of undulatory swimming in the American alligator. Copeia 4 , 839 - 843 (1984).
- Frolich, L. M. & Biewener, A. A. Kinematic and electromyographic analisys of the functional role of the body axis during terrestrial and aquatic locomotion in the salamander Ambystoma tigrinum. J. Exp. Biol. 162 , 107-130 (1992).
- Lauder G. V., Flammang B. E. & Alben S. Passive robotic models of propulsion by the bodies and caudal fins of fish. Integr. Comp. Biol. 52 , 576–587 (2012).
- Cavin, L. et al. Vertebrate assemblages from the early Late Cretaceous of southeastern Morocco: an overview. J. Afr. Earth Sci. 57 , 391- 412 (2010). 28. Meunier, L. M. V. & Larsson, H. C. E. Revision and phylogenetic affinities of Elosuchus (Crocodyliformes). Zool. J. Linnean Soc. 179 , 169-200 (2017). 29. Amiot, R. et al. Oxygen and carbon isotope compositions of middle Cretaceous vertebrates from North Africa and Brazil: ecological and environmental significance. Paleogeogr. Paleoclimatol Paleoecol. 297 , 439-451 (2010).
- Brusatte, S. L. Dinosaur Paleobiology (John Wiley & Sons, 2012).
- Taquet, P. The African cousins of the European iguanodontids. in Bernissart dinosaurs and Early Cretaceous terrestrial ecosystems (ed. Godefroit, P.) 283 – 291 (Indiana University Press, 20 12 ). Supplementary Information is available in the online version of the paper. Acknowledgements We thank Mhamed Azroal, Hamid Azroal, and M’Barek Fouadassi for assistance in the field. Ammar Aït Ha is thanked for help in preparing the fossils. The Moroccan Ministry of Mines, Energy and Sustainable Development is thanked for providing fieldwork permits. We thank Fabio Manucci for helpful discussions about the flesh reconstruction of Spinosaurus. This research was supported by a National Geographic Society grant to NI [CP-143R- 17 ], a National Geographic Emerging Explorer Grant to NI, contributions from the Board of Advisors of the University of Detroit Mercy to NI, a Jurassic Foundation grant to MF, a Paleontological Society grant to MF, an Explorers club grant to MF, as well as financial support from the Lokschuppen Rosenheim, the Museo di Storia Naturale di Milano, Joachim Pfauntsch, and Alessandro Lania.
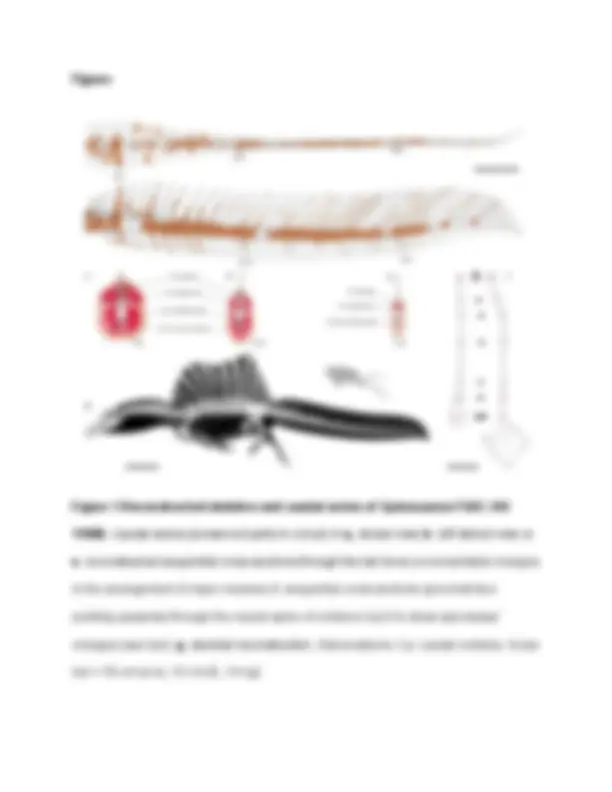
Figures Figure 1 Reconstructed skeleton and caudal series of Spinosaurus FSAC-KK
11888. Caudal series (preserved parts in colour) in a, dorsal view; b , left lateral view; c - e , reconstructed sequential cross sections through the tail show proximal/distal changes in the arrangement of major muscles; f , sequential cross sections (proximal face pointing upwards) through the neural spine of vertebra Ca23 to show apicobasal changes (see text). g , skeletal reconstruction. Abbreviations: Ca: caudal vertebra. Scale bar = 50 cm (a-e), 10 cm (f), 1m (g).
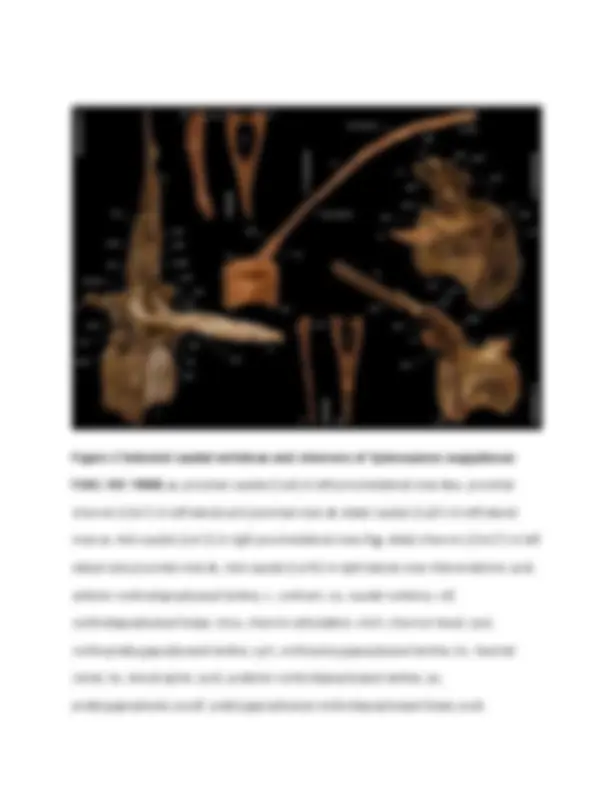
Figure 2 Selected caudal vertebrae and chevrons of Spinosaurus aegyptiacus FSAC-KK 11888. a , proximal caudal (Ca4) in left proximolateral view. b-c , proximal chevron (Chv7) in left lateral and proximal view. d , distal caudal (Ca31) in left lateral view. e , mid-caudal (Ca12) in right proximolateral view. f-g , distal chevron (Chv27) in left lateral and proximal view. h , mid-caudal (Ca16) in right lateral view. Abbreviations: acdl, anterior centrodiapophyseal lamina; c, centrum; ca, caudal vertebra; cdf, centrodiapophyseal fossa; chva, chevron articulation; chvh, chevron head; cpol, centropostzygapophyseal lamina; cprl, centroprezygapophyseal lamina; hc, haemal canal; ns, neural spine; pcdl, posterior centrodiapophyseal lamina; po, postzygapophysis; pocdf, postzygapophyseal centrodiapophyseal fossa; podl,
Fig. 3 : Thrust and efficiency of six different tail shapes during swimming. a, The Spinosaurus plastic tail shape attached to the robotic driving shaft in the water flume. The water is flowing from left to right at 10 cm/s. The coordinate system is provided in the top right. With reference to the tail, positive x forces (Fx) are generated anterior (or upstream), positive y forces (Fy) in the right lateral direction, and positive x forces (Fz)
in the ventral direction. b, Mean thrust and c, mean efficiency generated by tail shapes during robotically-controlled swimming. All tails were scaled to the same total length of 20 cm (Supplementary Information Fig. 4). The control tail was cut as a rectangular shape with the same surface area as the scaled Spinosaurus tail (63 cm 2 ). Details of the experimental setup are given in the methods. Raw thrust and efficiency data for each tail, including mean and standard error, are provided in Supplementary Data Table
- Swimming motions of the Spinosaurus tail can be found in Supplementary Videos 3-
Methods Excavation The Cretaceous Kem Kem beds of Morocco crop out along an extensive escarpment near the Moroccan-Algerian border region 7. After the accidental discovery and partial excavation by a local collector in 2008, part of a single skeleton (FSAC-KK 11888), subsequently deposited at the Faculté des Sciences of Casablanca University (FSAC), was recovered, published and designated as the neotype 7. A multi-institutional collaborative project in the years 2015-2019, led by NI, resulted in four joint expeditions to the neotype site. Detailed and careful exploration of the debris around the site, as well as a systematic and complex excavation of the unexcavated portion of the fossiliferous layer of the Zrigat hill, led to the recovery of many additional elements of the neotype skeleton (Extended Data Figure 1, Extended Data Figure 2). A detailed description of the new material, as well as the geological context, is included in the Supplemental Information. The supplemental information also includes details on a full-body flesh reconstruction based on FSAC-KK 11888. Experimental testing of tail shape swimming performance.
was tested N=5 times, except for the Spinosaurus tail which was tested N=5 times on two different days for a total of N=10 tests. Output data can be found in Supplementary Information Table 1. Thrust and efficiency for each tail shape were calculated using standard fluid dynamic equations as in our previous research 36 , 37. Mean thrust force ( Fx ) is calculated directly from transducer output from the Fx channel, and we accounted for transducer rotation resulting from the pitch motion to provide the force component directed upstream (positive thrust). Propulsive efficiency is calculated as the ratio of the thrust coefficient (𝐶𝑇 = 2 𝐹𝑥/𝜌𝑈^2 𝑐𝑠) to the power coefficient (𝐶𝑝 = 2 𝑃/𝜌𝑈^3 𝑐𝑠) where 𝜌 is the fluid density, U swimming velocity, c foil chord, and s the tail span. Effectively, this metric assesses the extent to which input power is translated into thrust. Osteohistological analysis. The aim of the osteohistological analysis was to determine if the remains assigned to FSAC-KK 11888 belong to a single individual rather than a chimaeric association of juvenile and adult individuals preserved in the same location and at the same horizon. The analysis was based on five selected elements. The primary assumption is that should histological details suggest that all five elements represent the same ontogenetic stage then they are more likely to represent one rather than multiple individuals. By contrast, should these elements exhibit two, or more, distinct ontogenetic stages this would point to the presence of multiple individuals of one, or perhaps several, taxa, all fortuitously preserved at a single location during a single depositional event38, 39,
The following elements were sectioned: the right femur; the left fibula; one rib; and two neural spines. All specimens were sectioned prior to preparation, in order to ensure that no outer layers
of the compact cortex were accidentally removed. In the case of the neural spines the apical portion was sectioned. Thin sectioning followed standard protocol 41. The thin sections have a thickness of 50- 70 microns and were analyzed with a petrographic microscope, Leica DM 2500 P. Digital images were captured using a ProgRes Cfscan camera. Only continuous lines were counted as lines of arrested growth (LAGs). Annuli were interpreted as a single year, following Lee and O’Connor 42. Retrocalculation, following the method proposed by Horner & Padian 43 , was applied to determine the likely number of missing LAGs, eroded through remodelling of the bone. In the case of the neural spines, only the width of the inner-most zone was used to retrocalculate the missing LAGs, because the shape of the section could not be approximated to a circular outline. The calculation of the major and minor axis used for the retrocalculation was performed in ImageJ 44. Results of the histological analysis are included in Supplemental Information.
- Lauder G. V., Anderson E. J., Tangorra J. & Madden P. G. A. Fish biorobotics: kinematics and hydrodynamics of self-propulsion. Journal of Experimental Biology 210 , 2767- 2780 (2007).
- Lauder G. V. et al. Robotic models for studying undulatory locomotion in fishes. Marine Technology Society Journal 45 , 41-55 (2011).
- Quinn, D. B., Lauder, G. V. & Smits, A. J. Maximizing the efficiency of a flexible propulsor using experimental optimization. Journal of Fluid Mechanics 767 , 430- 448 (2015).