Download Personality Stability in Late Adulthood: Genetic and Environmental Influences and more Study notes Statistics in PDF only on Docsity!
Personality Stability in Late Adulthood: A
Behavioral Genetic Analysis
Wendy Johnson, Matt McGue, and Robert F. Krueger
University of Minnesota–Twin Cities
ABSTRACT A sample of 833 twins from the Minnesota Twin Study of Adult Development and Aging completed the Multidimensional Personality Questionnaire (MPQ; Tellegen, 1982) twice, averaging 59. (sd 5 9.7) years of age at first and 64.4 (sd 5 10.2) years of age at second testing (average retest interval 5.0 years, sd 5 2.36, range 1.0–10.4 years). Both means and standard deviations of scale scores were extremely stable from first to second testing. In addition, sample participants tended to retain their rank order on the scales (average r 5 .76 across scales). Bivariate biometric analyses showed that the genetic influences on most of the scale scores were almost perfectly correlated across the two waves (range .95 to 1.00). The nonshared environmental influences were also highly correlated across the two waves (range .53 to .73). Models spec- ifying identical variance components at the two time points and fixing the genetic correlation to 1.00 provided improved fit. The results suggest that the high stability of personality in later adulthood has a strong genetic foundation, supplemented by stability of environmental effects.
A substantial body of evidence has accumulated for the stability of personality in adulthood. Such evidence is especially strong for the consistency of an individual’s relative position in a group (Roberts & DelVecchio, 2000). Consistency of this type, known as differential
The Minnesota Twin Study of Adult Development and Aging was supported by National Institute of Aging Grant #AG 06886. Robert F. Krueger was supported by National Institute of Mental Health Grant #MH 65137. Correspondence concerning this article should be addressed to Wendy Johnson, De- partment of Psychology, University of Minnesota, 75 East River Road, Minneapolis, MN 55455. E-mail john4350@tc.umn.edu.
Journal of Personality 73:2, April 2005 r Blackwell Publishing 2005 DOI: 10.1111/j.1467-6494.2005.00319.x
stability (Caspi & Roberts, 2001), is generally measured by a corre- lation coefficient. Such coefficients have been estimated for a variety of self-report personality questionnaires, ages, and retest intervals (e.g. Lubinski, Schmidt, & Benbow, 1996; McCrae & Costa, 1990; McGue, Bacon, & Lykken, 1993), but also for personality ratings by clinicians, spouses, and acquaintances (e.g. Conley, 1985; Costa & McCrae, 1988). In addition, there is strong evidence for the con- stancy of absolute levels of personality measures over time (Caspi & Roberts, 2001; Costa & McCrae, 1988). Roberts and DelVecchio (2000) concluded that the evidence to date suggests that relative sta- bility of personality over a period of several years increases with age, with correlations going from .31 in childhood to .64 around age 30, and leveling off at .74 for the ages 50 to 70. Stability of personality also decreases as the time interval between observations increases, yet impressive levels of stability have been observed over lengthy time spans in adulthood. Though 30-day re- test reliabilities for well-constructed scales can range from .80 to. (reflecting the measurement capacity of the instrument; Tellegen, 1982), retest stabilities across spans on the order of 25 years can re- main in the range of .60 to .70 (Costa & McCrae, 1992b; Helson & Klohnen, 1998), with higher stabilities (on the order of .65 to .80) reported for shorter time intervals of 6 to 12 years (Costa & McCrae, 1988, 1992b; Viken, Rose, & Koskenvuo, 1994). For adults under age 30, comparable stabilities for 6- to 10-year time intervals are only slightly lower, ranging from .48 to .60 (McGue et al., 1993; Viken et al., 1994). The particular aspect of personality measured appears to have little effect on the extent of stability, nor do sex or method of measurement (Roberts & DelVecchio, 2000). That genes contribute to personality throughout the lifespan is also very well established. Loehlin and Nichols’s (1976) study of al- most 800 pairs of adolescent twins and dozens of personality traits has stood the test of time, having been replicated many times using a variety of personality measurements and sample populations across a broad range of ages. Bouchard and Loehlin (2001) summarized the findings of 25 years of investigations as showing that 40–60% of in- dividual differences in personality can be attributed to genetic vari- ation. McCrae et al. (2000) have used this consistency of results across samples and many age ranges, construed in light of temperament as a constitutional predisposition to personality qualities, to argue that personality traits, even at the facet level, are genetically controlled
524 Johnson, McGue, & Krueger
circumstances rather than as changes in characteristics intrinsic to the individual such as personality. To address the gap, they describe the ways in which genetic and environmental influences, psycholog- ical function, and habitual person-environment transactions can contribute to stability. Very generally stated, these same processes can also contribute to change in the form of genetic influences on developmental processes, responses to environmental circumstances that reflect genetic as well as environmental influences, observational learning, learning generalizations, and learning from others’ descrip- tions of ourselves. They thus see both personality stability and change as the results of an ongoing series of transactions between genetic and environmental influences, which both have elements of stability and change. This conception also should be tested. In behavior genetic studies, environmental influences are generally categorized as either shared or nonshared. Shared environmental influences are those that contribute to similarity among reared-to- gether relatives. Frequently given examples of such influences refer to experiences of children, such as growing up with a common set of parents who provide a certain level of socioeconomic status, values such as religion, and common experiences such as travel, presence of alcoholism, or provision of a college education. But the concept of shared environmental influences can be extended to adults as well. It is easy to conceive of these kinds of experiences as having lifelong influences, and there is evidence for the existence of such effects for some of them. The process of measuring the magnitude of shared environmental influences does nothing to identify the specific influ- ences operating, so conceptual plausibility of this sort is all that is necessary to justify their quantification in this manner. Environmen- tal influences can be shared by siblings in adulthood as well. For example, siblings may continue to associate with each other fre- quently in adulthood, sharing holidays and care for elderly parents, among many other things. Thus, in adults, shared environmental influences include both lifelong effects of shared childhood experi- ences and the effects of currently shared experiences. The concept of shared environmental influence has had little effect on the concep- tion of personality emergence, stability, and change, however, be- cause most studies have shown little evidence of it for personality (Bouchard & Loehlin, 2001). Nonshared environmental influences are those that contribute to differences between siblings, and the same sort of discussion applies.
526 Johnson, McGue, & Krueger
In childhood, experiences such as having different schoolteachers, participating in different sports or other leisure activities, and dif- ferential parental treatment would qualify, and these experiences could have effects that persist into adulthood. In addition, in adult- hood, siblings typically have many different experiences involving spouses, jobs, and financial security (again among many other things). Nonshared environmental influences are rarely measured directly. Rather, they are measured as residual variance, which means that the term in which they are included always includes ran- dom error of measurement and often includes systematic method bias as well. Thus we would tend to expect relatively little relation- ship between nonshared environmental influences over time, due, in part, to measurement error and, in part, to changes in nonshared environmental circumstances over time. This means that one way to evaluate some of the implications of Roberts and Caspi’s (2003) and McCrae et al.’s (2000) conceptions of stability and change in per- sonality is to use a longitudinal twin study. When it is carried out in longitudinal form, the twin study method that serves as the cornerstone of behavioral genetic analysis can offer important contributions to our understanding of the nature of both the observed stability of and change in personality over time. In particular, such studies can address the extent to which genetic and environmental influences change with time by estimating the amounts of variance attributable to genetic and environmental in- fluences at varying time points. They can also address the extent to which genetic and environmental influences contribute to stability and change by estimating the extent to which the same genetic and environmental influences remain in place over time and new ones come into play. Loehlin (1992) reviewed longitudinal behavior ge- netic studies of variation in personality from childhood into adult- hood in order to summarize the evidence for various kinds of and changes in genetic influence. Though most of the studies he reviewed did not address genetic and environmental sources of personality stability and change directly, he concluded that although there are significant sources of genetic influence on personality change in childhood, after that, genetic influences contribute primarily to per- sonality stability rather than to change. This conclusion may have been driven by the relative lack of change in adulthood to evaluate. Genetic and environmental sources of stability of or change in personality in adulthood have been addressed specifically in just a
Personality Stability 527
adulthood, nor are we aware of any study that has investigated spe- cifically the question of environmental as well as genetic influences on stability over time. It is important to examine the stability of more specific personality traits as well as the broad traits such as Extraversion and Neuroticism because these broad traits are man- ifested in various specific ways, and stability at the level of the broad traits could mask change at the specific level. For example, both sociability and adventurousness are often considered to be aspects of Extraversion. A person, however, could change from being adven- turous but not particularly sociable to being sociable but not par- ticularly adventurous without changing in overall level of Extraversion. It is important to estimate the extent of environmen- tal as well as genetic influences on stability of personality because they can help us to distinguish between McCrae et al.’s (2000) con- ception of both stability of and change in personality as primarily genetic in origin and Roberts and Caspi’s (2003) perspective of a larger role for the environment in personality stability. Thus, the purpose of the current study was to investigate the ex- tent and sources of stability of and change in personality as meas- ured by the primary trait scales of the MPQ in late adulthood. We made use of the MPQ because it was designed to measure 11 specific personality traits with no item overlap among its primary scales (Tellegen, 1982). It therefore measures specific aspects of personality with high fidelity. Nevertheless, the primary scales of the MPQ cover a broad range of traits, showing good bandwidth. Thus, the MPQ combines fidelity and bandwidth in the measurement of personality, making it especially well suited to a comprehensive investigation of the genetic and environmental sources of personality stability. The MPQ is a self-report personality inventory that was con- structed by an exploratory factor-analytic process with the goal of developing relatively pure indexes of trait dispositions within normal personality. The result was 11 relatively independent primary trait scales. In factor analyses, the primary scales load on three higher- order factors, Positive Emotionality, Negative Emotionality, and Constraint, which relate to neurobiological mechanisms and map onto constructs of emotion and temperament. That is, Positive and Negative Emotionality incorporate temperamental dispositions to- ward those two emotions and are linked to brain motive systems underlying the appetitive-approach and defensive-withdrawal be- haviors (Derryberry & Reed, 1994; Lang, 1995; Sutton & Davidson,
Personality Stability 529
1997). Constraint incorporates traits related to behavioral restraint and reversed impulsivity, which has been characterized in terms of low brain arousal and reduced conditionability (Zuckerman, 1991). The primary MPQ scales have high internal consistency and 30-day retest reliabilities, and they cover a range of relatively psy- chometrically pure traits encompassing temperament, behavioral regulation, and interpersonal and imaginative styles. The MPQ’s combination of high reliability, construct independence, broad cov- erage of the personality domain, and neurobiological referents make it a particularly good choice for investigation of the extent and sources of personality stability. Table 1 briefly characterizes the MPQ scales. Positive Emotionality reflects the tendency to be actively and pleasurably engaged with one’s social and work environments; it is similar to the Extraversion dimension of the Eysenck Personality Questionnaire (EPQ; Eysenck, 1960) and the Five-Factor Model (FFM; McCrae & Costa, 1997) and to the Ascendance factor of the California Psychological Inventory (CPI; Gough & Bradley, 1996). The primary scales loading on positive emotionality reflect agency and communion; they are Wellbeing, Social Potency, Achievement, and Social Closeness. Negative Emotionality is characterized by perceptions of the world as threatening, problematic, and distress- ing; it is similar to Neuroticism as defined by many personality measures. Its primary scales reflect neurosis, estrangement, and confrontation; they are Stress Reactivity, Alienation, and Aggres- sion. Constraint is marked by the tendency to be restrained (impul- siveness reversed), conventional, and careful (sensation-seeking reversed). It is related to the Conscientiousness factor in the FFM (Church, 1994). The primary MPQ scales loading on Con- straint are Control, Harm Avoidance, and Traditionalism. The 11th primary MPQ trait scale, Absorption, reflects emotional responsive- ness to sensory input and loads on both Positive and Negative Emotionality. Specifically, in this study, we addressed three issues in a longitu- dinal, population-based Minnesota sample of primarily older-adult twins. First, we assessed the extent of personality stability and change, given measurements at two time points. Second, we esti- mated the genetic and environmental influences on the personality scale scores at both time points. Third, we investigated the extent to which the same genetic and environmental influences were involved
530 Johnson, McGue, & Krueger
at the two time points. We expected considerable stability of per- sonality, as well as a substantial genetic contribution to stability across time, as anticipated in the conceptions of the sources of per- sonality stability and change of both McCrae et al. (2000) and Roberts and Caspi (2003). To the extent that we also observed sub- stantial environmental contributions to stability across time, our re- sults would tend to provide greater support for Roberts and Caspi’s conception.
METHOD
Sample
The study sample was drawn from participants in the Minnesota Twin Study of Adult Development and Aging, a population-based registry in- cluding 901 intact monozygotic and same-sex dizygotic twin pairs born in the state of Minnesota. Most were born between 1904 and 1934, but 29% were born between 1936 and 1955, and one pair was born in 1897. This study has been described previously (Finkel & McGue, 1994). Briefly, participants were identified from Minnesota State Health Department birth records and recruited using state marriage and motor vehicle records; the initial participation rate was approximately 60%, likely due to the 4–5-hour length of the in-home assessment, which included a series of cognitive, personality, and lifestyle measures. Zygosity was determined using a five-item questionnaire that has been shown to provide over 95% accuracy (Lykken, Bouchard, McGue, & Tellegen, 1990). Twins were assigned as Twin 1 and Twin 2 by order of birth. A total of 1,123 participants (including 378 members of complete male twin pairs, 556 members of complete female twin pairs, and 189 twins whose co-twins did not provide data) completed the MPQ in the early 1990’s. In addition, 833 of these individuals (118 members of complete male MZ twin pairs, 106 members of complete male DZ twin pairs, 266 members of complete female MZ twin pairs, 168 members of complete female DZ twin pairs, and 90 male twins and 85 female twins whose co- twins did not provide data) completed the MPQ again, on average 5. years later (SD 2.4, range 1.0–10.4). Participants averaged 59.4 years of age (SD 9.7, range 27.0–92.0) at first and 64.4 (SD 10.2, range 32.2–98.7) at second administration. Though the full age range was rather large, the majority of participants were between ages 50 and 70 at first testing; less than 18% of participants were under age 50, and less than 10% were over age 70.
532 Johnson, McGue, & Krueger
Analysis of Mean-Level Personality Stability
To assess mean-level stability of our personality measures, we fit a hier- archical linear model (Bryk & Raudenbush, 1992), with overall mean scale score and retest interval as fixed effects and twin pair and initial score as random effects, to each set of MPQ scale scores. The use of in- terval of testing was necessary because the testing interval was reasonably evenly distributed over the range from 1.5 to 9 years. We made use of the hierarchical linear modeling technique rather than analysis of variance so that the estimates of statistical precision resulting from our models would be correctly adjusted both for the variable retest interval and for the correlations between scale scores within twin pairs.
Age and Sex Adjustment
Twins share a common age and, in our sample, a common sex. Thus age and sex effects act to increase twin similarity (McGue & Bouchard, 1984). To correct for these effects in the biometric analysis of the twin data, we adjusted the personality scores at the two time points by subtracting the sex-appropriate mean from each individual’s score and regressing out the linear component of age at each time point. In doing so, we did not norm the residuals to unit variance as we wished to be able to identify factors that contribute to variability in whatever personality change we observed.
Biometric Modeling
Our quantitative genetic model is based on the assumption that the ob- served phenotypic variance (Vp) is a linear additive function of genetic (A) and shared (C) and nonshared (E) environmental variance, respec- tively. Symbolically,
Vp¼ A þ C þ E:
Under this model, the nonshared environmental variance represents residual variance not explained by either of the other two sources. The nonshared environmental component also includes variance attributable to measurement error. Genetic variance can be additive in the sense that if multiple genes influence the trait, they do so independently of each other. It can also be nonadditive, reflecting dominance and other polygenic ef- fects. We note that there is evidence for nonadditive effects on person- ality, but little evidence for shared environmental effects (Bouchard & Loehlin, 2001; Finkel & McGue, 1997; Tellegen et al., 1988). For additive genetic effects, the expected covariance between any two members of a twin pair as a function of the variance components given above can be
Personality Stability 533
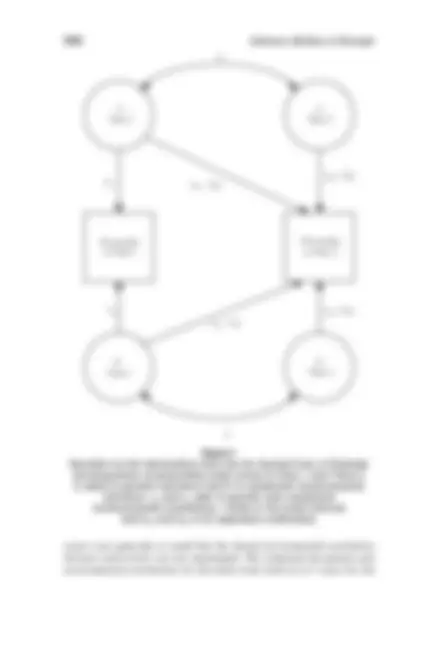
replicate), but little evidence for shared environmental effects (Tellegen et al., 1988; Bouchard & Loehlin, 2001). We began our biometric analyses by using Mx (Neale, 1997) to fit bivariate Cholesky models (ACE models) estimating additive genetic (termed the ‘‘A’’ parameter), and shared or common (termed the ‘‘C’’ parameter) and nonshared (termed the ‘‘E’’ parameter) environmental contributions to variance in and covariance between the age- and sex- adjusted MPQ personality scale scores at the two time points (referred to as Time 1 and Time 2). Because we had a substantial number of broken twin pairs in our sample, we made use of the raw data option in Mx so that the unmatched twins could contribute information about the vari- ances at the two time points. The path coefficients in the Cholesky models we fit were partial regression estimates, with the genetic and environ- mental path estimates to Time 1 stated directly. The effects of genetic and environmental influences on Time 2 personality, however, were decom- posed into those specific to Time 2 and those in common with Time 1. We explicitly made accommodation for the length of the retest interval by allowing it to moderate the decomposition of the genetic and environ- mental influences on Time 2 personality (a copy of the Mx script we used to do so can be obtained from the first author upon request). We derived two pieces of information from the Cholesky decomposi- tion. Figure 1 depicts the information we computed. First, we estimated the correlation between the genetic effects on personality at Time 1 and the genetic effects on personality at Time 2. This is known as the genetic correlation. The genetic correlation is defined as the square root of the proportion of the total heritability of the Time 2 scale score that can be explained by genetic effects at Time 1. Thus, when the genetic correlation is 1.00, all of the genetic influences are common to the two times. This also means that there is no room for genetic influence on whatever change has taken place over the interval. When the genetic correlation is 0.00, completely different sets of genetic influences act to affect individual dif- ferences at the two ages, and change could be genetic or environmental in origin to any degree. In other words, the genetic correlation reflects the extent to which the genetic effects overlap at the two times, regardless of their relative contribution to phenotypic variance. A similar interpreta- tion can be given to correlations of environmental influences, and we made the analogous calculation to estimate the nonshared environmental correlation between scale scores at Times 1 and 2. Thus, if stability in personality were primarily genetic in origin, we would expect high genetic correlations and very low environmental correlations. If stability in per- sonality reflected transactions between genes and environments, we would expect substantial correlations at both the genetic and environmental levels. The shared environmental influences on the personality scale
Personality Stability 535
scores were generally so small that the shared environmental correlation between scale scores was not meaningful. We computed the genetic and environmental correlations for the mean retest interval of 5 years for the
Personality at Time 1
Personality at Time 2
A Time 1
E Time 1
E Time 2
ra
r (^) e
a 22 + ba t
e 22 + b (^) e t
a (^11)
e 11 e 21 − bet
a 21 − b (^) a t
A Time 2
Figure 1 Heuristic for the information that can be derived from a Cholesky decomposition of personality scale scores at Time 1 and Time 2. A refers to genetic influence and E to nonshared environmental influence. ra and re refer to genetic and nonshared environmental correlations. t refers to the retest interval and ba and b (^) e to its regression coefficients.
536 Johnson, McGue, & Krueger
sample included some people spanning a large age range, we also explicitly checked for effects in our data of age on extent of change prior to beginning our biometric analyses. We did this by regressing change in raw scale scores on age and retest interval for each scale. There were no significant effects.
Biometric Analyses
We show the twin intraclass correlations for the primary and higher- order MPQ scales at Time 1 and Time 2 in Table 3. Because the MZ
Table 2 Significance Levels and Standardized Effect Sizes for the Mean Difference in Multidimensional Personality Questionnaire Scale Scores Over Time
Scale
Mean Time 1
Mean Time 2
T-value of difference
Degrees of freedom
Prob- ability
Standardized effect size
Primary Well-being 19.85 20.05 .92 1900 .36. Social potency 8.21 7.69 � 2.61 1900 .01 �. Achievement 12.07 11.76 � 3.31 1900 .001 �. Social closeness 15.70 15.89 .83 1894 .27. Stress reactivity 8.95 8.88 .65 1885 .52 �. Alienation 1.97 1.94 .88 1883 .38 �. Aggression 2.83 2.45 .55 1880 .58 �. Control 16.84 16.75 .89 1877 .37 �. Harm avoidance 22.25 22.64 2.18 1874 .03. Traditionalism 20.24 21.20 2.31 1867 .02. Absorption 14.30 13.96 .74 1847 .46 �. Higher order Positive emotion
152.35 152.00 � 1.17 1847 .24 �.
Negative emotion
123.15 122.46 � 1.93 1845 .05 �.
Constraint 171.87 173.07 .08 1847 .94.
Note. Mean differences were tested using a hierarachical linear model measuring fixed effects of retest interval. Effect size is (mean Time 2 less mean Time 1)/pooled standard deviation. It is stated for the average retest interval of 5 years. We made no adjustment to the probability levels for the multiple tests.
538 Johnson, McGue, & Krueger
correlations generally exceeded twice the DZ correlations, genetic influence is suggested for all personality scales, with little, if any, shared environmental influence. The few exceptions to this, showing DZ correlations more than half the MZ correlations, tended to be the scales loading on the Constraint superfactor. Assortative mating could possibly explain the relatively high DZ correlations, and we note that the Constraint superfactor has, in fact, shown more as- sortative mating than the other superfactors (Lykken & Tellegen, 1993). Table 4 summarizes the retest correlations within person and across MZ and DZ twin pairs. The cross-twin correlations express the degree of relationship between one twin’s score at Time 1 and the co-twin’s score at time 2 and therefore reflect the degree of associ- ation between the twins over time. Within-person correlations
Table 3 Monozygotic (MZ) and Dizygotic (DZ) Intraclass Correlations at Time 1 and Time 2 for the Multidimensional Personality Questionnaire Scales
Scale
Time 1 Time 2
MZ (pairs 5 254)
DZ (pairs 5 213)
MZ (pairs 5 175)
DZ (pairs 5 130)
Primary Well-being .27 .06 .33. Social potency .54 .23 .52. Achievement .46 .15 .39. Social closeness .45 .21 .46. Stress reactivity .49 .16 .46. Alienation .38 .16 .39. Aggression .29 .12 .48. Control .40 .11 .31. Harm avoidance .34 .22 .31. Traditionalism .57 .34 .55. Absorption .40 .29 .44. Higher order Positive emotion .43 .11 .40. Negative emotion .42 .24 .50. Constraint .53 .32 .54.
Note. Standard errors are less than .07 for MZ’s and .09 for DZ’s.
Personality Stability 539
or decreasing over time and the proportions of variance that result- ed. We computed the moderation effects of the retest interval for both genetic and nonshared environmental influences for all the scales, though we show the effects of these parameters only when they were significant. Each set of statistics measuring genetic and environmental variance components of scale scores was quite con- sistent across time for all of the scales, with the largest variations
Table 5 Computed Values From the Bivariate Cholesky Decomposition of the Multidimensional Personality Questionnaire Scale Across Time
Scale
A C E Totals
Mean time
Range over time
T1 T2 T1 T2 T1 T2 T1 T2 r (^) a r (^) e r (^) a r (^) e
Primary Well-being 27 31 0 0 73 68 100 99 .95 .65 1.00–.87 N/A Social potency 53 42 2 4 45 49 100 95 1.00 .70 N/A N/A Achievement 44 37 0 1 56 62 100 100 .96 .67 N/A N/A Social closeness 46 43 0 0 54 54 100 97 1.00 .63 N/A N/A Stress reactivity 41 30 4 10 55 60 100 100 1.00 .69 N/A N/A Alienation 14 23 25 25 61 56 100 103 .97 .67 1.00–.89 N/A Aggression 29 29 0 0 71 55 100 84 1.00 .57 N/A N/A Control 39 30 0 1 61 72 100 103 1.00 .60 N/A N/A Harm avoidance 40 34 0 0 60 64 100 98 1.00 .53 N/A N/A Traditionalism 38 36 17 5 45 39 100 80 1.00 .58 N/A N/A Absorption 27 35 15 8 58 63 100 106 1.00 .68 N/A N/A Higher order Positive emotion 42 41 0 0 58 63 100 104 .97 .73 N/A N/A Negative emotion 33 45 9 2 58 51 100 98 1.00 .71 N/A N/A Constraint 42 44 7 7 51 47 100 98 .93 .64 N/A .75–.
Note. A refers to additive genetic variance for the scale, C to shared environmental variance, and E to non-shared environmental variance; T1 and T2 refer to Time 1 and Time 2; ra and re refer to the genetic and non-shared environmental correlations between variance attributions at Time 1 and Time 2. Mean time genetic and envi- ronmental correlations are the estimates based on the mean time interval of 5 years. The range of these correlations over time refers to the estimates over the observed time interval of 1.0–10.0 years. The range is given only when the effect of retest interval was significant at p 5 .02 or less. The variance components have been scaled to the Time 1 Total level to enable examination of both changes in raw variances and proportions of variance.
Personality Stability 541
being on the order of 10%. The magnitudes of the proportional in- fluences on the personality scale scores were very comparable to those reported in other studies (Bouchard & Loehlin, 2001), except for the genetic influence on Alienation. That is, excluding Alienation and restating the raw variance components as proportions of the totals, the estimated genetic influences ranged from 27% to 53%, and estimated shared environmental influences were generally neg- ligible, though Alienation, Traditionalism, and Absorption, in par- ticular, showed some shared environmental influence. We believe that the unusually low estimate of genetic influence on Alienation resulted because the variance in that scale was notably smaller than those of the other scales with the exception of Aggression, resulting in depression of the correlations due to restriction of range. The freely estimated genetic correlations (ra ) for the mean retest interval of 5.0 years ranged from .93 to 1.00. The estimated nonshared en- vironmental correlations (r (^) e ) were also both significant and substan- tial; they ranged from .53 to .73. For the range of significant retest intervals included in the sample, the nonshared environmental cor- relations did not drop below .49. The high genetic correlations and small amounts of mean level change in the data implied that there were no substantive genetic influences on change in personality over the time interval. It was possible, however, that the small mean level changes masked sub- stantial changes at the individual level that could have either genetic or environmental influences. To evaluate this possibility, we com- puted the MZ and DZ twin correlations for change in raw scale scores over time. The largest correlation was .24, and it was the only one that was significant. Thus, consistent with the genetic correla- tions we computed, there was little or no evidence for genetic influ- ence on change in personality over time for this sample of primarily older adults. In our tables, we present the results for the ACE models, in spite of the fact that, for some of the models, including a parameter es- timating nonadditive genetic influence instead of a parameter esti- mating shared environmental influence fit better. There is substantial evidence for the existence of such nonadditive genetic influences on MPQ scales (Finkel & McGue, 1997; Tellegen et al., 1988). Their estimation, however, is fraught with difficulty as they cannot be es- timated simultaneously with both additive and shared environmental effects, and the power to estimate them is very weak (Martin, Eaves,
542 Johnson, McGue, & Krueger