Download Neural Correlates of Visual Awareness: Role of Visibility & Masking and more Study Guides, Projects, Research Cognitive Neuroscience in PDF only on Docsity!
Consciousness: Neurophysiology and Visual Awareness in
S L Macknik and S Martinez-Conde, Barrow Neurological Institute, Phoenix, AZ, USA Published by Elsevier Ltd.
Consciousness as a Neurophysiological
Problem
Consciousness is the feeling of life’s experience. It is a difficult concept to define precisely, however, because the word ‘consciousness’ means different things in different contexts. For instance, one may say that we are unconscious during sleep, despite the fact that, while asleep, we sometimes experience powerful and highly salient dreams (these dreams fit the defini- tion of consciousness as stated in the first sentence above). Does this mean that it is possible to be conscious of our unconscious experiences (an obvious semantic contradiction)? Or does this mean that dreams during sleep are not really unconscious? Because of these and many other semantic difficulties, the theorist Francis Crick (who also described the structure of DNA with James Watson), and his collaborator Christof Koch, suggested that we set aside the semantics and avoid defining the term consciousness linguistically. Instead, we should strive to establish the neural correlates of experience, or the neural correlates of consciousness (NCC). By determining the NCC, we will eventually arrive at a neurophysiological definition of conscious- ness. Following from this proposal, the field is currently at work to define consciousness by its neurophysiologi- cal underpinnings. Today, thanks in large extent to Crick and Koch’s efforts to validate the neurobiological study of consciousness, most neuroscientists and cog- nitive scientists are receptive to, and familiar with, the concept that consciousness, as with any other brain function (sensory, motor, emotional, and/or cognitive), must have a neurobiological substrate. In studying the NCC, one of the main strategies has been to make use of the vast library of knowledge that neurobiologists have accumulated, for many decades, on the primate visual system. Therefore, our dis- cussion of the neurophysiological underpinnings of consciousness focuses on those studies conducted in the visual system. The visual system of primates and humans offers the most extensive library we have available on the anatomy, physiology, and psycho- physics of perceptual experience. The authors of this article have begun to research the nature of the neuro- biological substrate of consciousness by isolating the neural correlates of a very small facet of our con- scious experience: the experience of visibility, which is arguably the most basic function of the
visual system. By ‘visibility,’ we do not mean the entire process of vision, but simply whether a stimulus is visible, or not. In this sense, visibility is the beginning of (or the necessary condition for) visual perception, not the conclusion. Without visibility, the stimulus cannot have significance or meaning: we perceive sti- mulus attributes such as color, motion, and depth only if the stimulus is also visible. Thus, when the underpinnings of visibility are discovered, they will include at least some of the circuits that cause aware- ness. This does not imply that visibility circuits must come before other types of circuits. For instance, it may not be necessary that a stimulus be visible before other types of processing take place (i.e., the corneal reflex). Whereas an unconscious visual reflex such as the accommodation of the eye may be a type of visual experience, stimuli that invoke visual reflexes are not necessarily consciously visible. That is, we are not aware of them despite the fact that they produce an appropriate and automatic response. As far as we know, visibility may take place subsequently to the processing of other visual attributes, or all visual at- tributes may come together in parallel, or there may be a disunity of processing for different attributes (as suggested by Semir Zeki), or there could be a combi- nation of all these possibilities. Whatever the sequence of events that eventually leads to visual awareness, we cannot be aware of a visual stimulus, by our defini- tion, unless it is visible.
Requirements for Establishing the NCC
Let us assume that visual awareness is correlated to brain activity within specialized neural circuits, and that not all brain circuits maintain awareness. It follows that the neural activity that leads to reflex- ive or involuntary motor action may not correlate with awareness because it does not reside within awareness-causing neural circuits. By our proposal, a stimulus that has become just-noticeable, or just-visible, has already activated awareness-producing circuits. Thus, awareness is neither a final process in vision, nor a very low-level process, but it is instead an intermediate-level pro- cess. A less than just-noticeable stimulus may activate neurons within early visual areas, but within circuits that do not maintain awareness; therefore, the stimu- lus remains invisible. Varying the level of salience of a percept (for instance, by varying its brightness above and beyond the just-noticeable threshold) may equate to varying the level of visibility and awareness. The idea of consciousness as an intermediate-level process, limited to intermediate brain areas, was first
Consciousness: Neurophysiology and Visual Awareness in 105
proposed by Jackendoff. According to Jackendoff’s proposal, the ‘homunculus’ in the frontal lobe is probably not conscious: we do not have direct con- scious access to our thoughts, but only to images, sounds, speech, and other bodily feelings associated with intermediate brain representations. Jackendoff’s theory provides a solution for the classical problem of the infinite regress of consciousness: given that the top homunculus is not conscious, there is no need for infinite homunculi at higher and higher levels in the brain in order to read the output of the previous, conscious homunculus. Let us begin with the hypothesis that there is a minimal set of conditions necessary to achieve visibil- ity, in the form of a specific type (or types) of neural activity within a subset of brain circuits. This minimal set of conditions will not be met if the correct circuits have the wrong type of activity (too much activity, too little activity, sustained activity when transient activity is required, etc). Moreover, if the correct type of activity occurs, but solely within circuits that do not maintain awareness, visibility will also fail. Finding the conditions in which visibility fails is criti- cal to the research described here: although we do not yet know what the minimal set of conditions is, we can nevertheless systematically modify potentially important conditions to see if they result in stimulus invisibility. If so, the modified condition is potentially part of the minimal set. To establish the minimal set of conditions for visibility we need to answer at least four questions:
- What stimulus parameters are important to visi- bility?
- What types of neural activity best maintain visibil- ity (transient vs. sustained firing, rate codes, bursts of spikes, etc. – that is, what is the neural code for visibility)?
- What brain areas must be active to maintain visibility?
- What specific neural circuits within the relevant brain areas maintain visibility? We must also determine the set of standards that will allow us to conclude that any given brain area, or neural circuit within an area, is responsible for gen- erating a conscious experience. Parker and Newsome developed a ‘‘list of idealized criteria that should be fulfilled if we are to claim that some neuron or set of neurons plays a critical role in the generation of a perceptual event.’’ If one replaces the words ‘‘percep- tual event’’ with ‘‘conscious experience,’’ Parker and Newsome’s list can be used as an initial foundation for the neurophysiological requirements needed to establish whether any given neuron or brain circuit
may be the neural substrate of awareness. The list is as follows:
- The responses of the neurons and of the perceiving subject should be measured and analyzed in directly comparable ways.
- The neurons in question should signal relevant information when the organism is carrying out the chosen perceptual task. Thus, the neurons should have discernable features in their firing patterns in response to the different external stimuli that are presented to the observer during the task.
- Differences in the firing patterns of some set of the candidate neurons to different external stimuli should be sufficiently reliable in a statistical sense to account for, and be reconciled with, the preci- sion of the organism’s responses.
- Fluctuations in the firing of some set of the candi- date neurons to the repeated presentation of identi- cal external stimuli should be predictive of the observer’s judgment on individual stimulus presen- tations.
- Direct interference with the firing patterns of some set of the candidate neurons (e.g., by electrical or chemical stimulation) should lead to some form of measurable change in the perceptual responses of the subject at the moment that the relevant external stimulus is delivered.
- The firing patterns of the neurons in question should not be affected by the particular form of the motor response that the observer uses to indi- cate his or her percept.
- Temporary or permanent removal of all or part of the candidate set of neurons should lead to a measurable perceptual deficit, however slight or transient in nature. However, visual circuits that may pass muster with Parker and Newsome’s guidelines may nevertheless fail to maintain awareness, as explained below. To isolate awareness-maintaining processes, some addi- tional strategies and principles must be added to guide the search for the NCC. The first strategy concerns the use of illusions as the tool of choice to test whether a neural tissue main- tains awareness. Visual illusions, by definition, disso- ciate the subject’s perception of a stimulus from its physical reality. Thus, visual illusions are powerful devices in the search for the NCC, as they allow the differentiation of neural responses to the physical stimulus from neural responses that correlate to per- ception. Our brains ultimately construct our percep- tual experience, rather than reconstruct the physical world. Therefore, an awareness-maintaining circuit should express activity that matches the conscious
106 Consciousness: Neurophysiology and Visual Awareness in
Since there is no monoptic form of binocular rivalry, one cannot use binocular rivalry by itself to test the strength of interocular suppression. One could use binocular rivalry in tandem with a different stimulus, such as visual masking stimuli (described below), to test for the explicit representation and strength of interocular suppression. But in such a case, the role of the tissue in maintaining visibility and awareness would have been probed by the visual masking stimuli, thus obviating the need for bin- ocular rivalry stimuli. Because one must rely on non- binocular rivalry stimuli to determine the explicit representation and strength of interocular suppres- sion in a given area, it is not possible to unambigu- ously interpret the neural correlates of perceptual state using binocular rivalry alone. Visual masking is a type of visual illusion that has been used as a primary tool to isolate the NCC. Visual masking illusions come in different flavors, but in all of them a visible target (a visual stimulus, such as a rectangle), or some specific aspect of a visible target (for instance, the semantic content of a word dis- played visually), is rendered invisible by changing the context in which the target is presented, without actually modifying the physical properties of the tar- get itself (Figure 1). That is, the target becomes less visible due solely to its spatial and/or temporal con- text. Visual masking illusions allow us to examine the brain’s response to the same physical target under varying levels of visibility. All we need to do is meas- ure the perceptual and physiological effects of the target when it is visible versus invisible and we will determine many, if not all, of the conditions that cause visibility. Our visual masking studies have shown that binocu- lar neurons in areas V1 (the first stage in the visual hierarchy where information from the two eyes is combined) and V2 of humans and monkeys can inte- grate excitatory responses between the eyes. How- ever, these same neurons do not express interocular suppression between the eyes. That is, binocular neu- rons in V1 are largely binocular for excitation while nevertheless being monocular for suppression. In summary, most early binocular cells do not explicitly process interocular suppression, and so these neurons cannot process binocular rivalry explicitly. Thus, bin- ocular rivalry is an inappropriate stimulus to probe these visual areas for the NCC. This finding renders the results from binocular rivalry studies in the early visual system uinterpretable with respect to localizing the NCC: the fact that early visual areas are not correlated to awareness of binocular rivalry is equi- valent in significance to concluding that these areas are not correlated to auditory awareness.
We propose that, to test for explicit processing in neural tissue, one should use a visual illusion with two modes of operation: one mode to ensure that the tissue processes the stimulus explicitly, and one mode to test the correlation to awareness. In visual masking, the monoptic mode establishes that the neural tissue processes masking stimuli explicitly, and then the dichoptic mode can be used to probe the NCC. The third strategy involves controlling for the effects of attention when designing experiments to isolate the NCC. Attention is a process in which the magnitude of neural activity is either enhanced or suppressed by high-level cognitive mechanisms. Attention may increase or decrease the likelihood of awareness of a given visual stimulus. But attention is a process distinct from awareness itself, as shown by Phillip Merikle and others. For instance, low-level bottom-up highly salient stimuli (such as flickering lights) can lead to awareness and draw attention, even when the subject is actively attending to some other task, or not attending to anything (i.e., when the subject is asleep). Thus, atten- tion can manipulate awareness, but the opposite is also true. This double-dissociation suggests that the two
Target only
Timeline Percept
Time
Time
Time
Time
Off
On
Off
Off
On Mask only
Target and mask
Target and mask
On
Off
On
Off
On
Off
On
Simultaneous target and mask
Backward masking
Figure 1 Temporal sequencing of stimuli in backward visual masking. A target and/or mask presented alone, or in simulta- neous combination, is visible. When the mask is presented after the target with the appropriate timing (backward masking), the target is rendered invisible. Visual masking illusions provide a powerful tool for investigating the neural underpinnings of visibility and invisibility.
108 Consciousness: Neurophysiology and Visual Awareness in
processes are mediated by separate brain circuits. It follows that in experiments to isolate the NCC, if the subject is conducting a task that requires attention to the stimulus of interest, then attentional and awareness processes may be confounded. Therefore, experiments to isolate the NCC should control for the effects of attention. If experimental manipulation of attentional state affects the magnitude of neural response, then the neural mechanism of interest may not be related to awareness, but instead to attention. Therefore, we add the following three criteria to Parker and Newsome’s list:
- The candidate neurons should be tested with an illusion that allows dissociation between the physical stimulus and its perception. If the candi- date set of neurons is capable of maintaining awareness, the neural responses should match the subjective percept, rather than the objective physical reality of the stimulus.
- The candidate neurons must explicitly pro- cess the type of information or stimulus used to test them.
- The responses of the neurons, and of the perceiving subject, should be measured with experimental controls for the effect of attention.
Techniques to Study the Neurophysiology
of Consciousness
The search for the NCC requires the localization of circuits in the brain that are sufficient to maintain awareness. To this end, brain areas have been sought within the ascending visual hierarchy that correlate, or more importantly, fail to correlate, with visual perception. For instance, the circuits of the brain that are critical to the visibility of targets must be circuits whose activity is suppressed during target invisibility due to visual masking. A corollary: if we identify circuits in which the target response is not suppressed during masking, we can rule out those circuits as sufficient to maintaining visual awareness. This section discusses the techniques used to identify parts of the brain whose activity correlates with vis- ibility and visual awareness. Table 1 compiles some of the neurophysiological techniques used in awareness research, as well as their strengths and weaknesses. All of these techniques may be paired with psychophysical measures of awareness. No single technique is perfect, but the combination of single-unit electrophysiology with anatomical analysis, fMRI, macroscopic optical recording, and two-photon laser scanning microscopy (2PLSM) may be most powerful to discover the cortical circuits that main- tain visual awareness in vivo.
Single-Unit Recordings Accompanied by Anatomical Analysis David Hubel and Torsten Wiesel championed the combined use of single-unit recordings and anatomi- cal staining techniques to achieve high-resolution cortical maps. The tungsten microelectrode, invented by Hubel, was similar to the electrodes used in humans by neurosurgeons such as Wilder Penfield and Benja- min Libet, but it was about 100–1000 times smaller at the tip, so it could be used to discover meaningful, if limited (on the scale of tens of microns), functional patterns within brain areas. Hubel and Wiesel com- bined electrode penetrations that were angled perpen- dicularly, obliquely, and tangentially to the cortical surface, with histological staining techniques to re- construct the electrode tracks in postmortem tissue. Clever application of these techniques led to the first model (on the scale of tens to hundreds of microns) of the two-dimensional pattern of visual function across the surface of the primary visual cortex. They dis- cerned functional domains: repeating patterns of physiological circuits across the surface of the cortex, such as ocular dominance and stimulus orientation columns. They also determined general principles of structure and function of the cortical layers: this added a third dimension to their functional anatomi- cal circuit model. Ultimately, Hubel and Wiesel found that mapping the three-dimensional cortex with a one-dimensional device was too limiting. By combining their record- ings with anatomical staining techniques, such as the Nauta method, radioactive 2-deoxyglucose uptake, horseradish peroxidase staining, and radioactive amino acid pathway tracing, they were able to better relate their electrode tracks to the direct postmortem analysis of cortical and subcortical maps. Modern versions of these methods are used today to determine the physiological underpinnings of visibil- ity and awareness in anesthetized and awake monkeys (Figure 2) and humans.
Functional Magnetic Resonance Imaging fMRI is the magnetic imaging of blood flow across differential behavioral conditions. Magnetic reso- nance imaging (MRI) scanners allow the radiologist to view the density of protons in the various tissues of the body in three-dimensional space, which essen- tially results in a very high resolution X-ray like image of the internal structures of the body. Blood is one of the tissues that can be imaged in the MR scanner. The magnetic properties of blood change as a function of neural activity: neural activity causes neurons to use more oxygen from the blood, and oxygenated blood has different magnetic properties
Consciousness: Neurophysiology and Visual Awareness in 109
personal copy
than deoxygenated blood. Therefore, functional MR imaging of blood flow (fMRI) is particularly ame- nable to magnetic imaging analysis. The basis is the following: iron (deoxygenated hemoglobin) is mag- netic, whereas rust (oxygenated hemoglobin) is not magnetic. Thus, by isolating the parts of the brain in which blood changes its magnetic properties in corre- lation to a task, we can infer where the neurons of the brain have responded functionally. It was this line of reasoning that led Ogawa and colleagues to discover that BOLD signal can be used to map brain function noninvasively. fMRI has sparked a major revolution in the discov- ery and mapping of human cortical brain areas. How- ever, the method has relatively low spatial resolution (on the order of millimeters, rather than the micron resolution we enjoy with anatomical staining tech- niques). It also has low temporal resolution (the hemo- dynamic neural response function occurs over seconds, rather than the microsecond resolution we enjoy with electrophysiology). Figure 3 shows a map of many of the retinotopic visual areas that may be relevant to maintaining visual awareness. These visual areas can be probed with illusory stimuli, such as visual masking stimuli, to determine whether BOLD activity correlates with the subject’s perception (Figure 4).
Macroscopic Optical Imaging Lack of oxygen not only changes blood from nonmag- netic to magnetic, but also turns its color from red (rusty) to bluish (iron). The regions of the brain’s sur- face that are active during a given task remove more oxygen from the blood, and so these regions appear bluer than the regions where neurons are less active. Gary Blasdel and Guy Salama discovered that when red light was shone onto the exposed surface of the cortex, more red photons were absorbed by the active regions, as blue blood absorbs more red light than red blood. This decrease in reflectance can be measured with a video camera to map cortical function. fMRI and macroscopic optical recording of intrinsic signals have the same physical substrate (blood oxyge- nation level), and therefore the same slow temporal resolution. However, macroscopic optical recording enjoys a much higher spatial resolution than fMRI: the resolution of the capillary network just below the brain’s surface is tens to hundreds of microns, depend- ing on the tissue. Figure 5 illustrates how macroscopic optical maps may be used to examine area V1 responses to stan- dard visual stimuli. Figure 6 shows an optical image
Target mask
Time
Forward maskingBackward masking
100 ms
The standing wave of invisibility illusion
50 ms a
b
M M M
10 �
LGN T T Single-unit
0 100 200 Time (ms)
300 400
100
200 Spikes/s
300
0
Figure 2 The electrophysiology of visual masking. (a) The standing wave of invisibility (SWI): a temporal sequence of target and mask presentations that results in very powerful masking of the target. (b) A single-neuron recording from the lateral genicu- late nucleus (LGN) of a monkey. The stimulus presentation layout on the monkey’s computer monitor is illustrated in the inset of each row. The cross is the monkey’s fixation point. The target (T) is centered over the receptive field of the LGN neuron and color-coded in black (actual presentation color was white on a black background). The mask (M) is color-coded in purple (actual color same as target) and is presented at four different distances from the target (one distance per row). The physiological traces are represented in black (when the target was presented alone), purple (when the mask was presented alone), and blue (when the target and mask were presented together in the SWI condition). Notice that when the mask is near the target, the target response during the SWI condition (blue) is lower than the target response when presented alone; this is the neural correlate of visual mask- ing. As the mask is moved away from the target, the target response during the SWI condition recovers its magnitude, just as the target becomes more visible perceptually. Reproduced from Macknik SL, Martinez-Conde S, and Haglund MM (2000) The role of spatiotemporal edges invisibility and visual masking. Proceedings of the National Academy of Sciences of the United States of America 97: 7556–7560, with permission from the National Academy of Sciences of the United States of America.
Consciousness: Neurophysiology and Visual Awareness in 111
of V1 responses to visual masking illusions, such as the standing wave of invisibility.
Two-Photon Laser Scanning Microscopy One of the great problems with standard light micros- copy is that the light source is inevitably brighter than the tissue of interest, which drastically decreases the quality of the imaging. To make matters worse, the images are also contaminated by light coming from sources outside of the plane of focus. The invention of fluorescence microscopy was a major leap forward for microscopic analysis: it allowed for the illumina- tion of tissue with one color of light (the excitation wavelength) while imaging only the fluoresced photons of a different color (the emission wavelength). Thus, fluorescence microscopes filter out the photons outside the emission wavelength, which improves the quality of the imaging greatly. The main problem with fluores- cent imaging in biological tissues is that fluorescence, by its quantum mechanical nature, turns a high-energy (bluish) photon into a lower energy (reddish) photon. Because brain tissue tends to absorb blue photons, one must use extremely high-powered light sources to image tissues more than �100 microns deep; this carries the risk of burning the brain. In 1990, Denk et al. used two-photon fluorescence excitation to work around these problems. The idea behind the two-photon quantum effect is that a fluo- rescent molecule, which has an excitation wavelength of, say, 400 nm, will fluoresce a photon when excited
by two photons of 800 nm (which each have exactly half the energy of a single 400 nm photon). So the same fluorescent dye will fluoresce a greenish emis- sion photon after receiving a single bluish (400 nm) excitation photon, or two simultaneous infrared (800 nm) photons. The advantage is that the brain is relatively transparent to infrared light (as compared with blue light) and so one can image much deeper and with a much lower power light source (i.e., infra- red) than when using a blue light source. The illumi- nation method is moreover inherently focused on a given focal plane because the two infrared photons must land on the same exact fluorescent dye molecule within femptoseconds of each other to result in the emission of a green photon. Any excitation (infrared) light that is scattered by irrelevant tissue outside the focal point does not cause noise in the imaging (it is simply lost), and so only the single point of focus (in three dimensions) of the laser will generate fluores- cence. Because of these features, 2PLSM solves all of the problems above: it combines fluorescence (which allows one to filter out noise from the light source) with the ability to use an infrared light source to image deeply within the cortex (on the order of 1 mm), and furthermore to maintain an optical resolution that is roughly the size of a single photon. 2PLSM has not yet been used to examine cir- cuits that are critical to visibility or consciousness. However, it is only a matter of time before this method is employed to determine the microscopic neural inter- actions that are the fundamental basis of awareness.
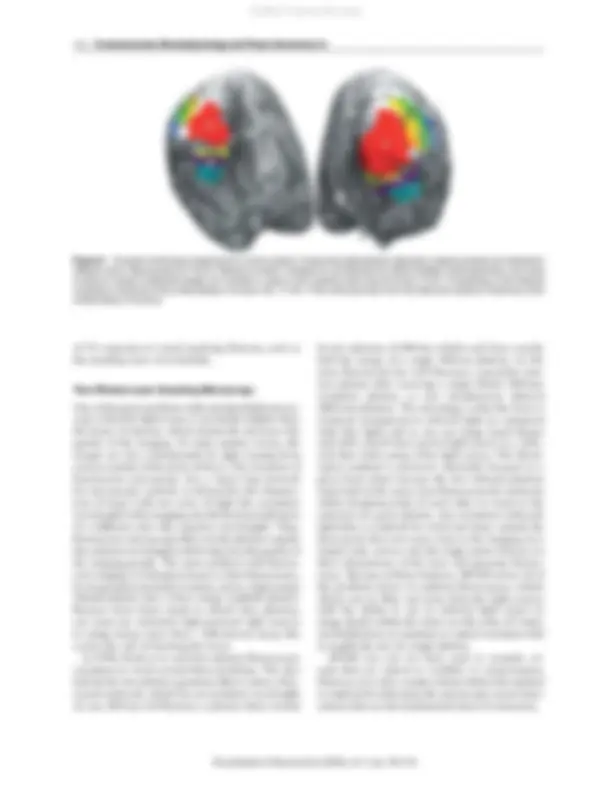
Figure 3 Example of retinotopy mapping from a human subject. Visual areas delineated by retinotopic mapping analysis are indicated in different colors. Reproduced from Tse Pu, Martinez-Conde S, Schlegel AA, and Macknik SL (2005) Visibility, visual awareness, and visual masking of simple unattended targets are confined to areas in the occipital cortex beyond human V1/V2. Proceedings of the National Academy of Sciences of the United States of America 102: 17178–17183, with permission from the National Academy of Sciences of the United States of America.
112 Consciousness: Neurophysiology and Visual Awareness in
responses) matched perception, making area V3 the lower boundary in the visual hierarchy for the proces- sing of visual awareness. Moreover, we found that only areas within the occipital lobe correlate with awareness of simple unattended targets. This finding provided us with the first experimental evidence of an upper boundary (the edge of the occipital lobe) for the processing and maintenance of visual aware- ness. These results localized visual awareness of sim- ple unattended targets to a relatively small part of the brain, between area V3 and the edge of the occipital lobe. Future research will more precisely localize the specific circuits within this region that maintain awareness. The results support Jackendoff’s theory that con- sciousness is an intermediate-level process, and not a low- or high-level process. However, it remains possible (contrary to Jackendoff’s proposal) that higher brain areas, such as the frontal lobes, maintain awareness of
higher cognitive functions and that only sensory awareness is maintained at intermediate levels.
Future Challenges
The field of the neurobiology of consciousness is at its very incipient stage. We and others have begun to narrow down the potential brain regions or circuits that may maintain various types of sensory aware- ness. However, we are far from having a solid under- standing of the NCC. Once the NCC are localized, a bigger challenge will be to determine the specific nature of the underlying circuits that maintain aware- ness. This is further complicated by the possibility that there may be multiple different circuits that maintain awareness of different types of events. On the bright side, the field is converging on a set of techniques and methods that can be used to test neural tissue at multiple levels of resolution, both
1 cm
0.13� wide
0.64�
~2� ~2�
Figure 5 The optical image of a flickering bar. The left column represents the layout of the stimuli used to stimulate the cortical window imaged in the right column. Top: Image of the area V1 cortical intrinsic signal generated by a flickering bar 50 ms on and 100 ms off, 0.13� wide with an orientation of 132�. The imaged patch is 1 cm^2 and was approximately 10–12�^ below and to the left of the foveal representation and subtended about 4�^ of visual angle (as measured with microelectrode penetrations at each edge of the image) at the anterior-medial border of the operculum. The vertical meridian is parallel to the lower edge of the image; the fovea is to the right. Bottom: Image of the intrinsic signal generated by a flickering bar 0.64�^ wide in the same piece of cortex (the center of the bar was also shifted approximately 0.29�^ away from the fovea). Notice that the widened optical image of the bar has shifted in position and split into two edges, showing that edges generate much stronger signals that the interiors of objects at the level of V1. Reproduced from Macknik SL, Martinez-Conde S, and Haglund MM (2000) The role of spatiotemporal edges invisibility and visual masking. Proceedings of the National Academy of Sciences of the United States of America 97: 7556–7560, with permission from the National Academy of Sciences of the United States of America.
114 Consciousness: Neurophysiology and Visual Awareness in
invasively and noninvasively. Once these techniques are fully developed, long hours in the lab and clever experimental design will accomplish much of the remaining work.
See also: Attentional Networks; Awareness: Functional Imaging; Blindsight: Residual Vision; Coma and Other Pathological Disorders of Consciousness; Conscious- ness: Theories and Models; Consciousness: Theoretical and Computational Neuroscience; Consciousness: Neu- ral Basis of Conscious Experience; Contextual Interac- tions in Visual Perception; Contextual Interactions in Visual Processing.
Further Reading
Blasdel GG and Salama G (1986) Voltage-sensitive dyes reveal a modular organization in monkey striate cortex. Nature 321: 579–585. Crick FHC (1994) The Astonishing Hypothesis: The Scientific Search for the Soul. New York: Touchstone. Denk W, Strickler JH, and Webb WW (1990) Two-photon laser scanning fluorescence microscopy. Science 248: 73–76. Hubel DH and Wiesel TN (2005) Brain and Visual Perception: The Story of a 25-Year Collaboration. New York: Oxford University Press. Jackendoff R (1987) Consciousness and the Computational Mind. Cambridge, MA: MIT Press. Koch C (2004) The Quest for Consciousness: A Neurobiological Approach. CO: Roberts and Publishers.
Libet B (2006) Reflections on the interaction of the mind and brain. Progress in Neurobiology 78: 322–326. Macknik SL (2006) Visual masking approaches to visual aware- ness. Progress in Brain Research 155: 179–217. Macknik SL and Haglund MM (1999) Optical images of visible and invisible percepts in the primary visual cortex of primates. Proceedings of the National Academy of Sciences of the United States of America 96: 15208–15210. Macknik SL and Livingstone MS (1998) Neuronal correlates of visibility and invisibility in the primate visual system. Nature Neuroscience 1: 144–149. Macknik SL and Martinez-Conde S (2004) Dichoptic visual masking reveals that early binocular neurons exhibit weak interocular suppression: Implications for binocular vision and visual awareness. Journal of Cognitive Neuroscience 16: 1–11. Macknik SL and Martinez-Conde S (2004) The spatial and tempo- ral effects of lateral inhibitory networks and their relevance to the visibility of spatiotemporal edges. Neurocomputing 58–60C: 775–782. Macknik SL, Martinez-Conde S, and Haglund MM (2000) The role of spatiotemporal edges in visibility and visual masking. Proceedings of the National Academy of Sciences of the United States of America 97: 7556–7560. Malonek D and Grinvald A (1996) Interactions between electrical activity and cortical microcirculation revealed by imaging spec- troscopy – Implications for functional brain mapping. Science 272: 551–554. Martinez-Conde S (2006) Fixational eye movements in normal and pathological vision. Progress in Brain Research 154: 153–178. Martinez-Conde S, Macknik SL, and Hubel DH (2000) Microsaccadic eye movements and firing of single cells in
a b c
Stimulus
Response
Target
Mask
Mask
Target
Mask
Mask
Figure 6 Optical imaging of the standing wave of invisibility (SWI). (a) A visual target with a width of 0.12�^ and the correlated optical image. (b) Two masking stimuli presented without a target and the correlated response. (c) The SWI stimulus (target and masks were never presented on the display at the same time) and the correlated response. Notice that the image of the target in (c) is now missing, compared to the response seen in (a), despite the fact that both targets were physically identical. Reproduced from Macknik SL and Haglund MM (1999) Optical images of visible and invisible percepts in the primary visual cortex of primates. Proceedings of the National Academy of Sciences of the United States of America 96: 15208–15210, with permission from the National Academy of Sciences of the United States of America.
Consciousness: Neurophysiology and Visual Awareness in 115